
α2δ-4 and Cachd1 Proteins Are Regulators of Presynaptic Functions
 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Results
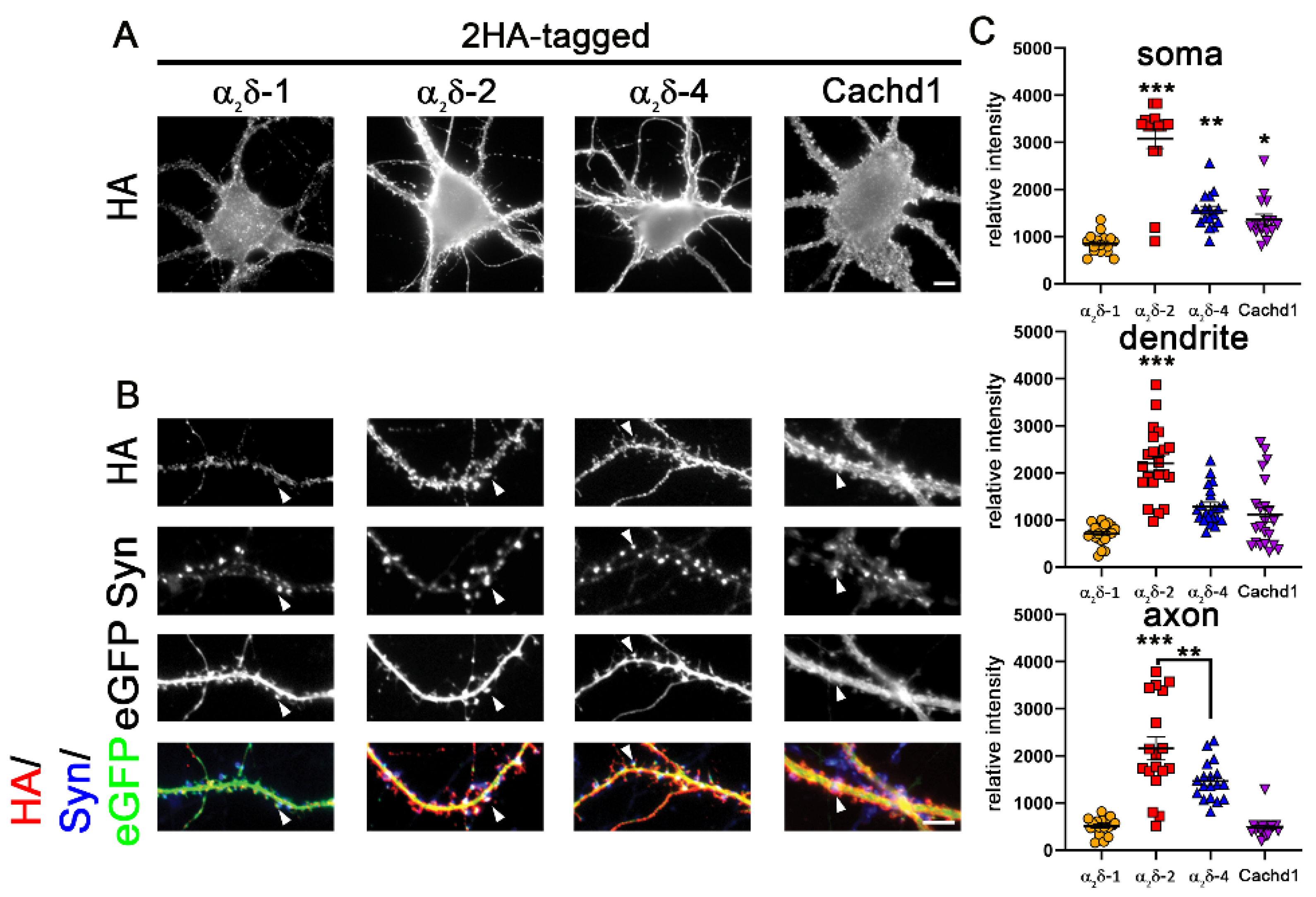
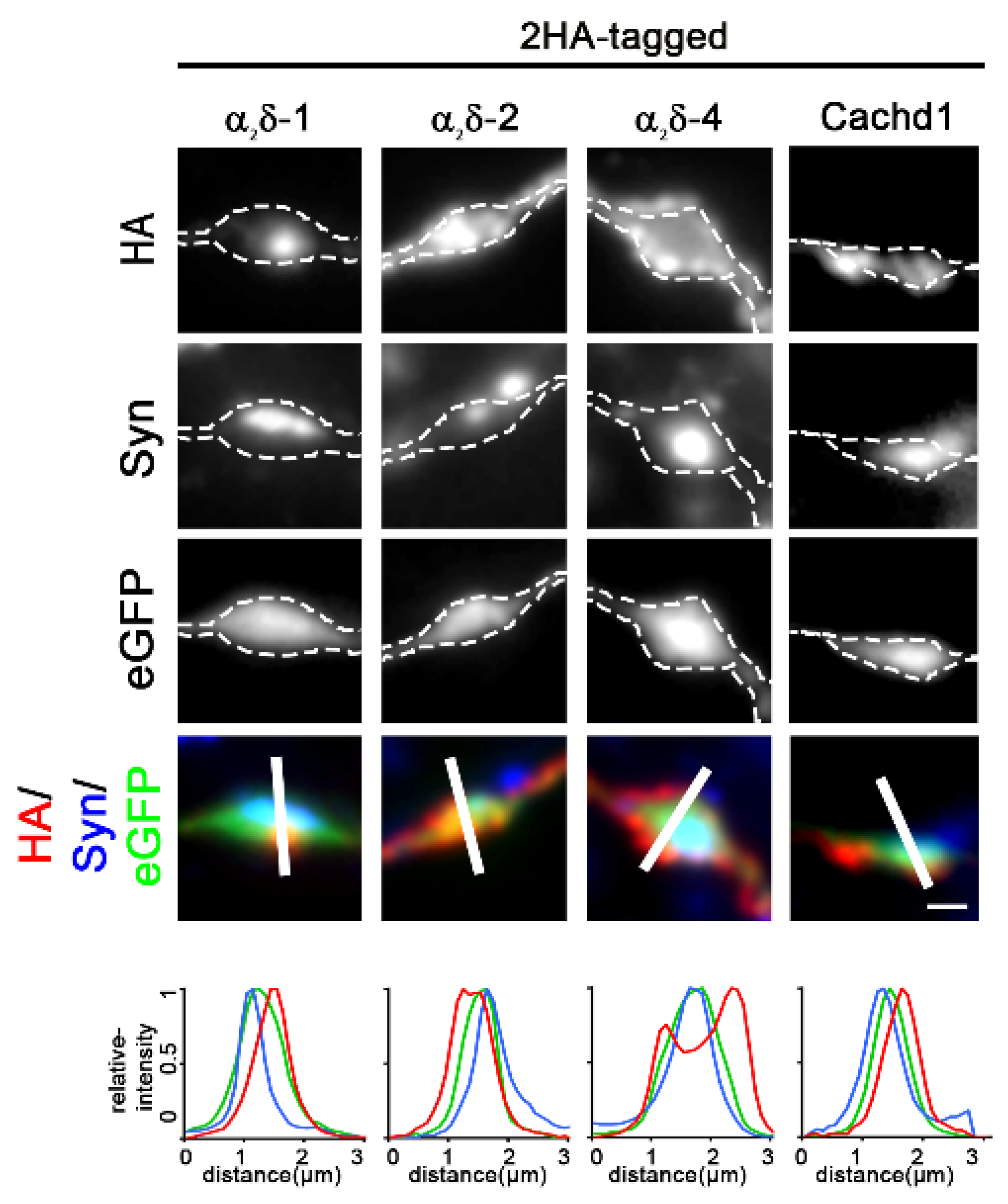
2.1. Epitope-Tagged α2δ-4 and Cachd1 Target to Presynaptic Terminals
2.2. α2δ-4 and Cachd1 Rescue Glutamatergic Synapse Formation
2.3. α2δ-4 and Cachd1 Restore Presynaptic Calcium Signaling in Triple Knockout Neurons
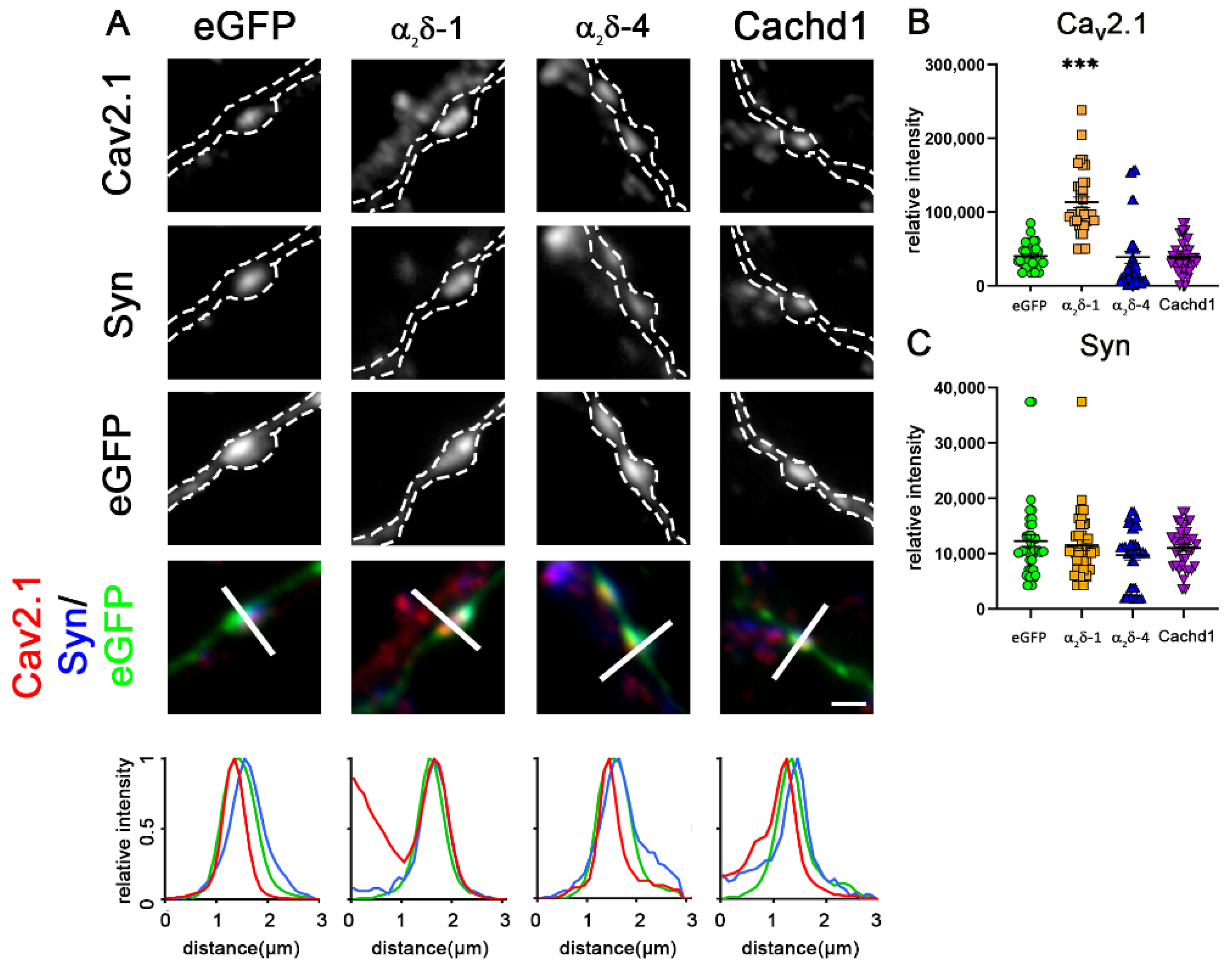
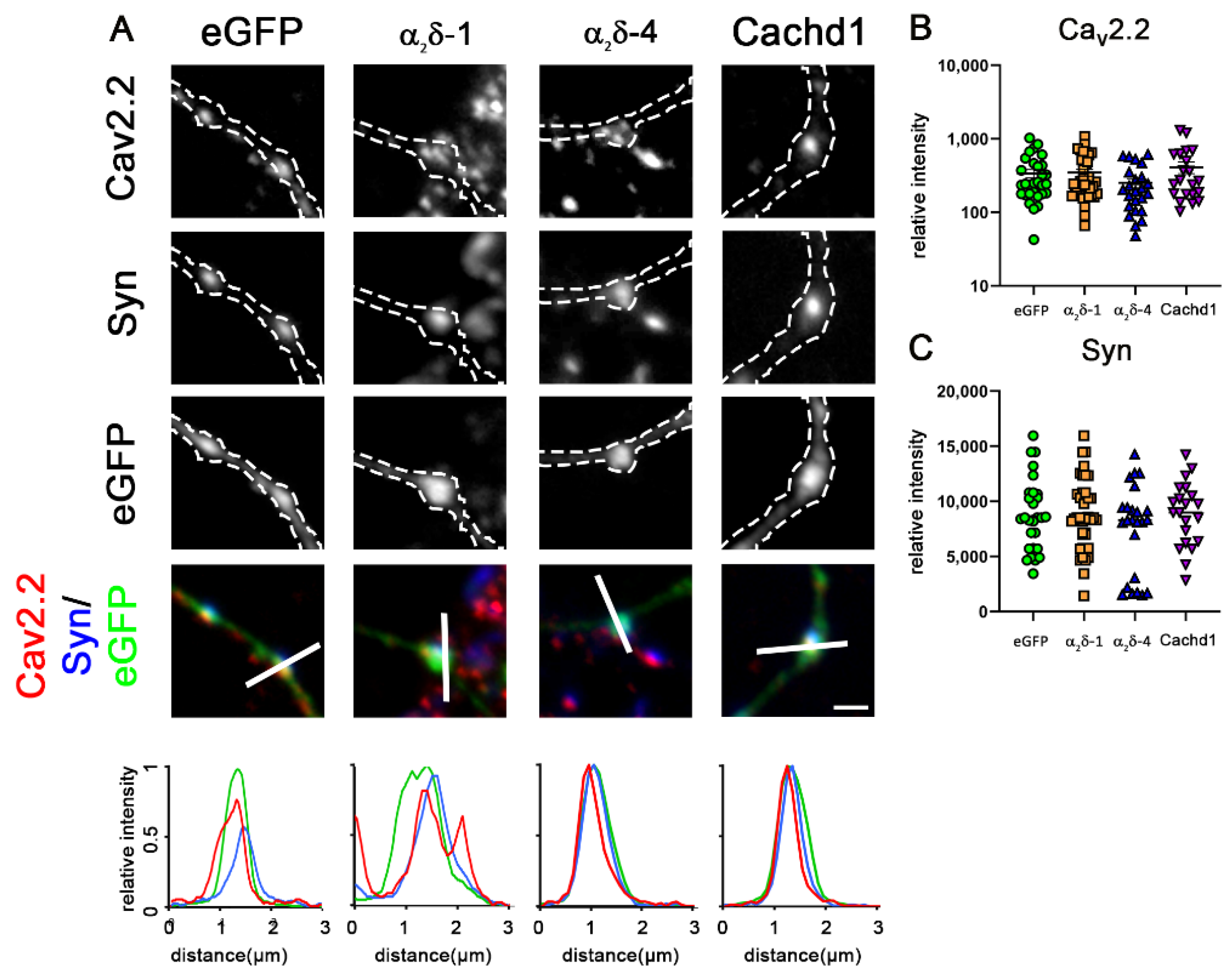
2.4. α2δ-4 and Cachd1 Do Not Increase Presynaptic CaV2.1 Abundance
2.5. α2δ-4 and Cachd1 Do Not Affect Glutamatergic Synapse Composition
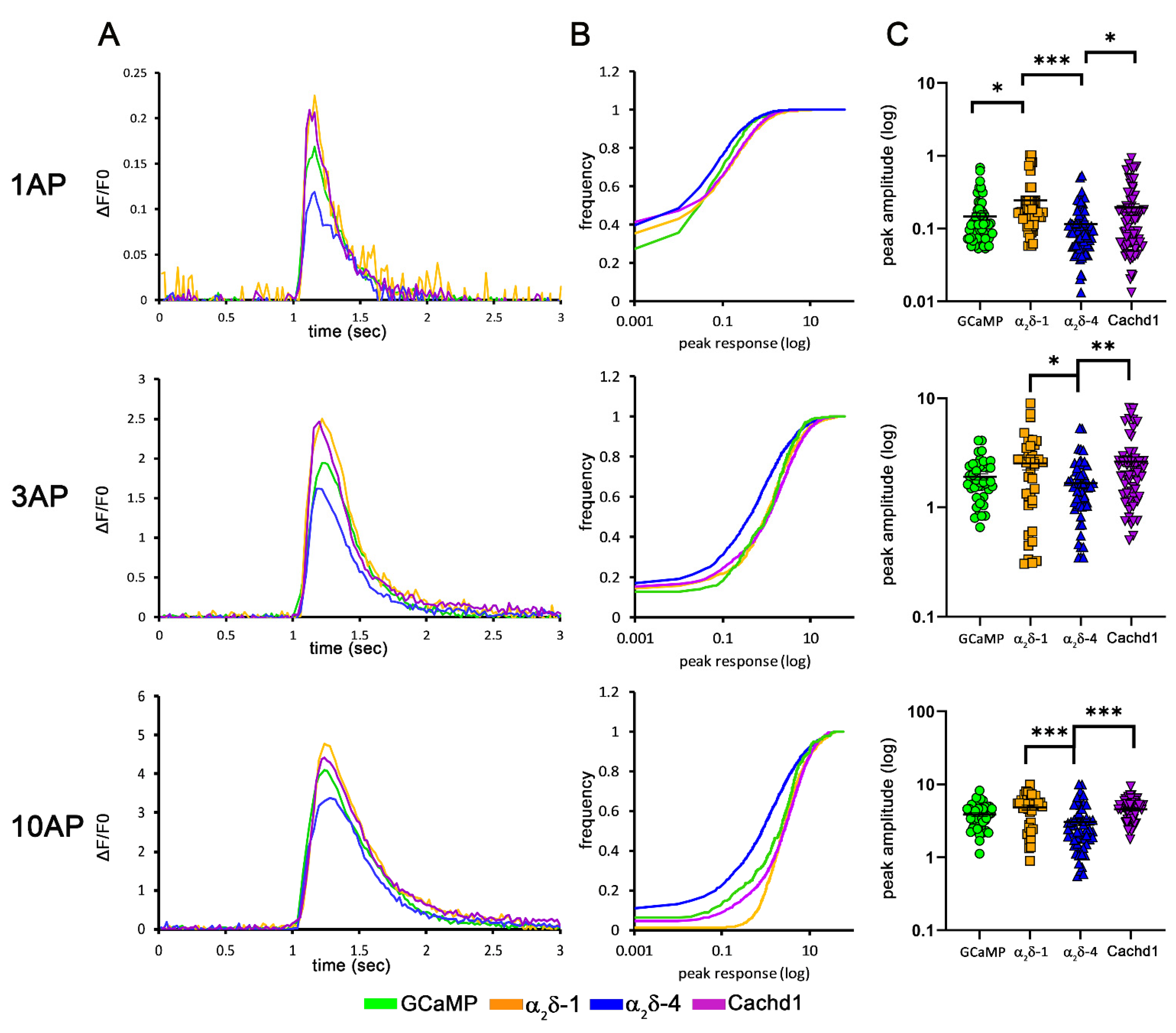
2.6. Differential Effects on Presynaptic Calcium Transients
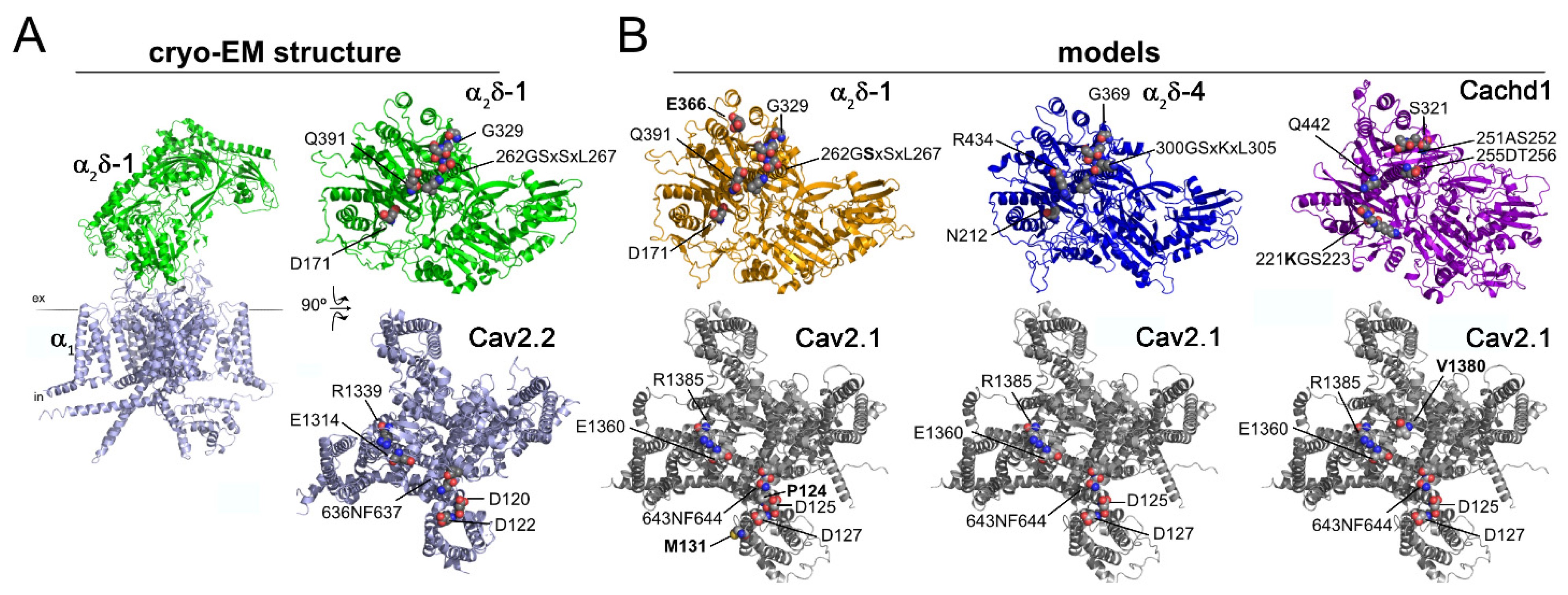
2.7. Structural Determinants for Protein–Protein Interactions of α2δ Subunits and Cachd1 with Pore-Forming α1 Subunits
3. Discussion
3.1. Functional Redundancy of α2δ Proteins
3.2. Differential Presynaptic Effects of α2δ-4 and Cachd1
3.3. Synaptic Roles of α2δ-4 and Cachd1
3.4. Conclusions
4. Materials and Methods
4.1. Breeding and Genotyping Procedures
4.1.1. Animals
4.1.2. Breeding and Genotyping of Mutant Mice
4.2. Cell Culture and Transfection Procedures
Primary Cultures of Hippocampal Neurons for Fluorescence Imaging
4.3. Expression Vectors and Cloning Procedures
4.4. High-Resolution Fluorescence Microscopy
Antibodies
4.5. Image Analysis and Quantification
4.5.1. Colocalization of Synaptic Proteins
4.5.2. Quantification of Fluorescent Clusters in Single Boutons
4.5.3. Quantification of Live Cell Surface Expression
4.5.4. Quantification Analysis
4.6. Calcium Imaging
Data Analysis
4.7. Homology Modeling
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dolphin, A.C. Calcium Channel Auxiliary α2δ and β Subunits: Trafficking and One Step Beyond. Nat. Rev. Neurosci. 2012, 13, 542–555. [Google Scholar] [CrossRef] [PubMed]
- Felix, R.; Gurnett, C.A.; de Waard, M.; Campbell, K.P. Dissection of Functional Domains of the Voltage-Dependent Ca2+ Channel Alpha2delta Subunit. J. Neurosci. 1997, 17, 6884–6891. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A. Voltage-Gated Calcium Channels. Cold Spring Harb. Perspect. Biol. 2011, 3, a003947. [Google Scholar] [CrossRef] [PubMed]
- Calderón-Rivera, A.; Andrade, A.; Hernández-Hernández, O.; González-Ramírez, R.; Sandoval, A.; Rivera, M.; Gomora, J.C.; Felix, R. Identification of a Disulfide Bridge Essential for Structure and Function of the Voltage-Gated Ca2+ Channel α2δ-1 Auxiliary Subunit. Cell Calcium 2012, 51, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Ablinger, C.; Geisler, S.M.; Stanika, R.I.; Klein, C.T.; Obermair, G.J. Neuronal α2δ Proteins and Brain Disorders. Pflügers Arch.-Eur. J. Physiol. 2020, 472, 845–863. [Google Scholar] [CrossRef]
- Andrade, A.; Brennecke, A.; Mallat, S.; Brown, J.; Gomez-Rivadeneira, J.; Czepiel, N.; Londrigan, L. Genetic Associations between Voltage-Gated Calcium Channels and Psychiatric Disorders. Int. J. Mol. Sci. 2019, 20, 3537. [Google Scholar] [CrossRef]
- Eroglu, C.; Allen, N.J.; Susman, M.W.; O’Rourke, N.A.; Park, C.Y.; Ozkan, E.; Chakraborty, C.; Mulinyawe, S.B.; Annis, D.S.; Huberman, A.D.; et al. Gabapentin Receptor Alpha2delta-1 Is a Neuronal Thrombospondin Receptor Responsible for Excitatory CNS Synaptogenesis. Cell 2009, 139, 380–392. [Google Scholar] [CrossRef]
- Risher, W.C.; Kim, N.; Koh, S.; Choi, J.-E.; Mitev, P.; Spence, E.F.; Pilaz, L.-J.; Wang, D.; Feng, G.; Silver, D.L.; et al. Thrombospondin Receptor α2δ-1 Promotes Synaptogenesis and Spinogenesis via Postsynaptic Rac1. J. Cell Biol. 2018, 217, 3747–3765. [Google Scholar] [CrossRef]
- Beeson, K.A.; Beeson, R.; Westbrook, G.L.; Schnell, E. α2δ-2 Protein Controls Structure and Function at the Cerebellar Climbing Fiber Synapse. J. Neurosci. 2020, 40, 2403–2415. [Google Scholar] [CrossRef]
- Fell, B.; Eckrich, S.; Blum, K.; Eckrich, T.; Hecker, D.; Obermair, G.J.; Münkner, S.; Flockerzi, V.; Schick, B.; Engel, J. α2δ2 Controls the Function and Trans-Synaptic Coupling of Cav1.3 Channels in Mouse Inner Hair Cells and Is Essential for Normal Hearing. J. Neurosci. 2016, 36, 11024–11036. [Google Scholar] [CrossRef] [Green Version]
- Geisler, S.; Schöpf, C.L.; Stanika, R.; Kalb, M.; Campiglio, M.; Repetto, D.; Traxler, L.; Missler, M.; Obermair, G.J. Presynaptic α2δ-2 Calcium Channel Subunits Regulate Postsynaptic GABAA Receptor Abundance and Axonal Wiring. J. Neurosci. 2019, 39, 2581–2605. [Google Scholar] [CrossRef] [PubMed]
- Pirone, A.; Kurt, S.; Zuccotti, A.; Rüttiger, L.; Pilz, P.; Brown, D.H.; Franz, C.; Schweizer, M.; Rust, M.B.; Rübsamen, R.; et al. α2δ3 Is Essential for Normal Structure and Function of Auditory Nerve Synapses and Is a Novel Candidate for Auditory Processing Disorders. J. Neurosci. 2014, 34, 434–445. [Google Scholar] [CrossRef] [PubMed]
- Caylor, R.C.; Jin, Y.; Ackley, B.D. The Caenorhabditis Elegans Voltage-Gated Calcium Channel Subunits UNC-2 and UNC-36 and the Calcium-Dependent Kinase UNC-43/CaMKII Regulate Neuromuscular Junction Morphology. Neural Dev. 2013, 8, 10. [Google Scholar] [CrossRef]
- Kurshan, P.T.; Oztan, A.; Schwarz, T.L. Presynaptic α2δ-3 Is Required for Synaptic Morphogenesis Independent of Its Ca2+-Channel Functions. Nat. Neurosci. 2009, 12, 1415–1423. [Google Scholar] [CrossRef] [PubMed]
- Schlick, B.; Flucher, B.E.; Obermair, G.J. Voltage-Activated Calcium Channel Expression Profiles in Mouse Brain and Cultured Hippocampal Neurons. Neuroscience 2010, 167, 786–798. [Google Scholar] [CrossRef]
- Kerov, V.; Laird, J.G.; Joiner, M.-L.; Knecht, S.; Soh, D.; Hagen, J.; Gardner, S.H.; Gutierrez, W.; Yoshimatsu, T.; Bhattarai, S.; et al. α2δ-4 Is Required for the Molecular and Structural Organization of Rod and Cone Photoreceptor Synapses. J. Neurosci. 2018, 38, 6145–6160. [Google Scholar] [CrossRef] [PubMed]
- Knoflach, D.; Kerov, V.; Sartori, S.B.; Obermair, G.J.; Schmuckermair, C.; Liu, X.; Sothilingam, V.; Garcia Garrido, M.; Baker, S.A.; Glösmann, M.; et al. Cav1.4 IT Mouse as Model for Vision Impairment in Human Congenital Stationary Night Blindness Type 2. Channels 2013, 7, 503–513. [Google Scholar] [CrossRef]
- Wang, Y.; Fehlhaber, K.E.; Sarria, I.; Cao, Y.; Ingram, N.T.; Guerrero-Given, D.; Throesch, B.; Baldwin, K.; Kamasawa, N.; Ohtsuka, T.; et al. The Auxiliary Calcium Channel Subunit α2δ4 Is Required for Axonal Elaboration, Synaptic Transmission, and Wiring of Rod Photoreceptors. Neuron 2017, 93, 1359–1374. [Google Scholar] [CrossRef]
- Wycisk, K.A.; Budde, B.; Feil, S.; Skosyrski, S.; Buzzi, F.; Neidhardt, J.; Glaus, E.; Nürnberg, P.; Ruether, K.; Berger, W. Structural and Functional Abnormalities of Retinal Ribbon Synapses Due to Cacna2d4 Mutation. Investig. Ophthalmolo. Vis. Sci. 2006, 47, 3523–3530. [Google Scholar] [CrossRef]
- van Loo, K.M.J.; Rummel, C.K.; Pitsch, J.; Müller, J.A.; Bikbaev, A.F.; Martinez-Chavez, E.; Blaess, S.; Dietrich, D.; Heine, M.; Becker, A.J.; et al. Calcium Channel Subunit α2δ4 Is Regulated by Early Growth Response 1 and Facilitates Epileptogenesis. J. Neurosci. 2019, 39, 3175–3187. [Google Scholar] [CrossRef] [Green Version]
- Klomp, A.; Omichi, R.; Iwasa, Y.; Smith, R.J.; Usachev, Y.M.; Russo, A.F.; Narayanan, N.S.; Lee, A. The Voltage-Gated Ca2+ Channel Subunit α2δ-4 Regulates Locomotor Behavior and Sensorimotor Gating in Mice. PLoS ONE 2022, 17, e0263197. [Google Scholar] [CrossRef] [PubMed]
- Cottrell, G.S.; Soubrane, C.H.; Hounshell, J.A.; Lin, H.; Owenson, V.; Rigby, M.; Cox, P.J.; Barker, B.S.; Ottolini, M.; Ince, S.; et al. CACHD1 Is an α2δ-Like Protein That Modulates CaV3 Voltage-Gated Calcium Channel Activity. J. Neurosci. 2018, 38, 9186–9201. [Google Scholar] [CrossRef] [PubMed]
- Stephens, G.J.; Cottrell, G.S. CACHD1: A New Activity-Modifying Protein for Voltage-Gated Calcium Channels. Channels 2019, 13, 120–123. [Google Scholar] [CrossRef] [PubMed]
- Dahimene, S.; Page, K.M.; Kadurin, I.; Ferron, L.; Ho, D.Y.; Powell, G.T.; Pratt, W.S.; Wilson, S.W.; Dolphin, A.C. The α2δ-like Protein Cachd1 Increases N-Type Calcium Currents and Cell Surface Expression and Competes with α2δ-1. Cell Rep. 2018, 25, 1610–1621. [Google Scholar] [CrossRef]
- Schöpf, C.L.; Ablinger, C.; Geisler, S.M.; Stanika, R.I.; Campiglio, M.; Kaufmann, W.A.; Nimmervoll, B.; Schlick, B.; Brockhaus, J.; Missler, M.; et al. Presynaptic α2δ Subunits Are Key Organizers of Glutamatergic Synapses. Proc. Natl. Acad. Sci. USA 2021, 118, e1920827118. [Google Scholar] [CrossRef]
- Obermair, G.J.; Szabo, Z.; Bourinet, E.; Flucher, B.E. Differential Targeting of the L-Type Ca2+ Channel Alpha 1C (CaV1.2) to Synaptic and Extrasynaptic Compartments in Hippocampal Neurons. Eur. J. Neurosci. 2004, 19, 2109–2122. [Google Scholar] [CrossRef]
- Stanika, R.; Campiglio, M.; Pinggera, A.; Lee, A.; Striessnig, J.; Flucher, B.E.; Obermair, G.J. Splice Variants of the CaV1.3 L-Type Calcium Channel Regulate Dendritic Spine Morphology. Sci. Rep. 2016, 6, 34528. [Google Scholar] [CrossRef]
- Brockhaus, J.; Schreitmüller, M.; Repetto, D.; Klatt, O.; Reissner, C.; Elmslie, K.; Heine, M.; Missler, M. α-Neurexins Together with α2δ-1 Auxiliary Subunits Regulate Ca2+ Influx through Cav2.1 Channels. J. Neurosci. 2018, 38, 8277–8294. [Google Scholar] [CrossRef]
- Scott, M.B.; Kammermeier, P.J. CaV2 Channel Subtype Expression in Rat Sympathetic Neurons Is Selectively Regulated by α2δ Subunits. Channels 2017, 11, 555–573. [Google Scholar] [CrossRef]
- Hoppa, M.B.; Lana, B.; Margas, W.; Dolphin, A.C.; Ryan, T.A. α2δ Expression Sets Presynaptic Calcium Channel Abundance and Release Probability. Nature 2012, 486, 122–125. [Google Scholar] [CrossRef] [Green Version]
- Geisler, S.M.; Benedetti, A.; Schöpf, C.L.; Schwarzer, C.; Stefanova, N.; Schwartz, A.; Obermair, G.J. Phenotypic Characterization and Brain Structure Analysis of Calcium Channel Subunit α2δ-2 Mutant (Ducky) and α2δ Double Knockout Mice. Front. Synaptic Neurosci. 2021, 13, 634412. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Yao, X.; Yan, N. Structure of Human Cav2.2 Channel Blocked by the Painkiller Ziconotide. Nature 2021, 596, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Held, R.G.; Liu, C.; Ma, K.; Ramsey, A.M.; Tarr, T.B.; de Nola, G.; Wang, S.S.H.; Wang, J.; van den Maagdenberg, A.M.J.M.; Schneider, T.; et al. Synapse and Active Zone Assembly in the Absence of Presynaptic Ca2+ Channels and Ca2+ Entry. Neuron 2020, 107, 667–683.e9. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.; Flodman, P.L.; Gargus, J.J.; Simon, M.T.; Verrell, K.; Haas, R.; Reiner, G.E.; Naviaux, R.; Osann, K.; Spence, M.A.; et al. Mitochondrial and Ion Channel Gene Alterations in Autism. Biochim. Biophys. Acta 2012, 1817, 1796–1802. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S.M.; Moran, J.L.; Fromer, M.; Ruderfer, D.; Solovieff, N.; Roussos, P.; O’Dushlaine, C.; Chambert, K.; Bergen, S.E.; Kähler, A.; et al. A Polygenic Burden of Rare Disruptive Mutations in Schizophrenia. Nature 2014, 506, 185–190. [Google Scholar] [CrossRef]
- van den Bossche, M.J.; Strazisar, M.; de Bruyne, S.; Bervoets, C.; Lenaerts, A.-S.; de Zutter, S.; Nordin, A.; Norrback, K.-F.; Goossens, D.; de Rijk, P.; et al. Identification of a CACNA2D4 Deletion in Late Onset Bipolar Disorder Patients and Implications for the Involvement of Voltage-Dependent Calcium Channels in Psychiatric Disorders. Am. J. Med. Genetics. Part B Neuropsychiatr. Genet. 2012, 159B, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Rudan Njavro, J.; Klotz, J.; Dislich, B.; Wanngren, J.; Shmueli, M.D.; Herber, J.; Kuhn, P.-H.; Kumar, R.; Koeglsperger, T.; Conrad, M.; et al. Mouse Brain Proteomics Establishes MDGA1 and CACHD1 as in Vivo Substrates of the Alzheimer Protease BACE1. FASEB J. 2020, 34, 2465–2482. [Google Scholar] [CrossRef]
- Moran, Y.; Zakon, H.H. The Evolution of the Four Subunits of Voltage-Gated Calcium Channels: Ancient Roots, Increasing Complexity, and Multiple Losses. Genome Biol. Evol. 2014, 6, 2210–2217. [Google Scholar] [CrossRef]
- Neely, G.G.; Hess, A.; Costigan, M.; Keene, A.C.; Goulas, S.; Langeslag, M.; Griffin, R.S.; Belfer, I.; Dai, F.; Smith, S.B.; et al. A Genome-Wide Drosophila Screen for Heat Nociception Identifies α2δ3 as an Evolutionarily Conserved Pain Gene. Cell 2010, 143, 628–638. [Google Scholar] [CrossRef]
- Brodbeck, J.; Davies, A.; Courtney, J.-M.; Meir, A.; Balaguero, N.; Canti, C.; Moss, F.J.; Page, K.M.; Pratt, W.S.; Hunt, S.P.; et al. The Ducky Mutation in Cacna2d2 Results in Altered Purkinje Cell Morphology and Is Associated with the Expression of a Truncated Alpha2Delta-2 Protein with Abnormal Function. J. Biol. Chem. 2002, 277, 7684–7693. [Google Scholar] [CrossRef] [Green Version]
- Barclay, J.; Balaguero, N.; Mione, M.; Ackerman, S.L.; Letts, V.A.; Brodbeck, J.; Canti, C.; Meir, A.; Page, K.M.; Kusumi, K.; et al. Ducky Mouse Phenotype of Epilepsy and Ataxia Is Associated with Mutations in the Cacna2d2 Gene and Decreased Calcium Channel Current in Cerebellar Purkinje Cells. J. Neurosci. 2001, 21, 6095–6104. [Google Scholar] [CrossRef] [PubMed]
- Kaech, S.; Banker, G. Culturing Hippocampal Neurons. Nat. Protoc. 2006, 1, 2406–2415. [Google Scholar] [CrossRef] [PubMed]
- Obermair, G.J.; Schlick, B.; Di Biase, V.; Subramanyam, P.; Gebhart, M.; Baumgartner, S.; Flucher, B.E. Reciprocal Interactions Regulate Targeting of Calcium Channel Beta Subunits and Membrane Expression of Alpha1 Subunits in Cultured Hippocampal Neurons. J. Biol. Chem. 2010, 285, 5776–5791. [Google Scholar] [CrossRef]
- Bacchi, N.; Messina, A.; Burtscher, V.; Dassi, E.; Provenzano, G.; Bozzi, Y.; Demontis, G.C.; Koschak, A.; Denti, M.A.; Casarosa, S. A New Splicing Isoform of Cacna2d4 Mimicking the Effects of c.2451insC Mutation in the Retina: Novel Molecular and Electrophysiological Insights. Investig. Ophthalmol. Vis. Sci. 2015, 56, 4846–4856. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating Signal Peptides from Transmembrane Regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Folci, A.; Steinberger, A.; Lee, B.; Stanika, R.; Scheruebel, S.; Campiglio, M.; Ramprecht, C.; Pelzmann, B.; Hell, J.W.; Obermair, G.J.; et al. Molecular Mimicking of C-Terminal Phosphorylation Tunes the Surface Dynamics of CaV1.2 Calcium Channels in Hippocampal Neurons. J. Biol. Chem. 2018, 293, 1040–1053. [Google Scholar] [CrossRef]
- Di Biase, V.; Flucher, B.E.; Obermair, G.J. Resolving Sub-Synaptic Compartments with Double Immunofluorescence Labeling In Hippocampal Neurons. J. Neurosci. Methods 2009, 176, 78–84. [Google Scholar] [CrossRef]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and Development of Coot. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef]
- Krissinel, E.; Henrick, K. Inference of Macromolecular Assemblies from Crystalline State. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Dilution | Source |
|---|---|---|
| Anti-HA | 1:100 (LIVE/A594) | Roche (Mannheim, Germany) (catalog #11867423001) |
| Anti-GABAARβ2/3 | 1:500 (A594) | Millipore (Darmstadt, Germany) (catalog #MAB341) |
| 1:250 (A350) | Millipore (catalog #MAB341) | |
| Anti-gephyrin | 1:2000 (A594) | Synaptic Systems (Göttingen, Germany) (catalog #147 021) |
| Anti-GLUR1 | 1:1000 (A594) | Upstate (Lake Placid, NY, USA) (catalog #06-306) |
| Anti-PSD-95 | 1:1000 (A594) | Thermo Fisher Scientific (Waltham, MA, USA) (catalog #MA1-045) |
| Anti-synapsin1 | 1:500 (A350) | Synaptic Systems (catalog #106 011) |
| Anti-vGLUT1 | 1:2000 (A350) | Synaptic Systems (catalog #135 002) |
| Anti-vGLUT1 | 1:500 (A594) | Synaptic Systems (catalog #135 511) |
| 1:250 (A350) | Synaptic Systems (catalog #135 511) | |
| Anti-vGAT | 1:500 (A350) | Synaptic Systems (catalog #131 002) |
| Anti-Cav2.1 | 1:2000 (A594) | Synaptic Systems (catalog #152203) |
| Anti-Cav2.2 | 1:2000 (A594) | Synaptic Systems (catalog #152313) |
| Goat anti-Mouse IgG, Alexa Fluor 350 | 1:500 | Thermo Fisher Scientific (catalog #A-21049) |
| Goat anti-Rabbit IgG, Alexa Fluor 350 | 1:500 | Thermo Fisher Scient (catalog #A-21068) |
| Goat anti-Mouse IgG, Alexa Fluor 594 | 1:4000 | Thermo Fisher Scientific (catalog #A-11032) |
| Goat anti-Rabbit IgG, Alexa Fluor 594 | 1:4000 | Thermo Fisher Scientific (catalog #A-11037) |
| Goat anti-Rat IgG, Alexa Fluor 594 | 1:4000 | Thermo Fisher Scientific (catalog #A-11007) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ablinger, C.; Eibl, C.; Geisler, S.M.; Campiglio, M.; Stephens, G.J.; Missler, M.; Obermair, G.J. α2δ-4 and Cachd1 Proteins Are Regulators of Presynaptic Functions. Int. J. Mol. Sci. 2022, 23, 9885. https://doi.org/10.3390/ijms23179885
Ablinger C, Eibl C, Geisler SM, Campiglio M, Stephens GJ, Missler M, Obermair GJ. α2δ-4 and Cachd1 Proteins Are Regulators of Presynaptic Functions. International Journal of Molecular Sciences. 2022; 23(17):9885. https://doi.org/10.3390/ijms23179885
Chicago/Turabian StyleAblinger, Cornelia, Clarissa Eibl, Stefanie M. Geisler, Marta Campiglio, Gary J. Stephens, Markus Missler, and Gerald J. Obermair. 2022. "α2δ-4 and Cachd1 Proteins Are Regulators of Presynaptic Functions" International Journal of Molecular Sciences 23, no. 17: 9885. https://doi.org/10.3390/ijms23179885