Dynamics of Cyanobacteria and Related Environmental Drivers in Freshwater Bodies Affected by Mitten Crab Culturing: A Study of Lake Guchenghu, China

Abstract
:1. Introduction
2. Materials and Methods
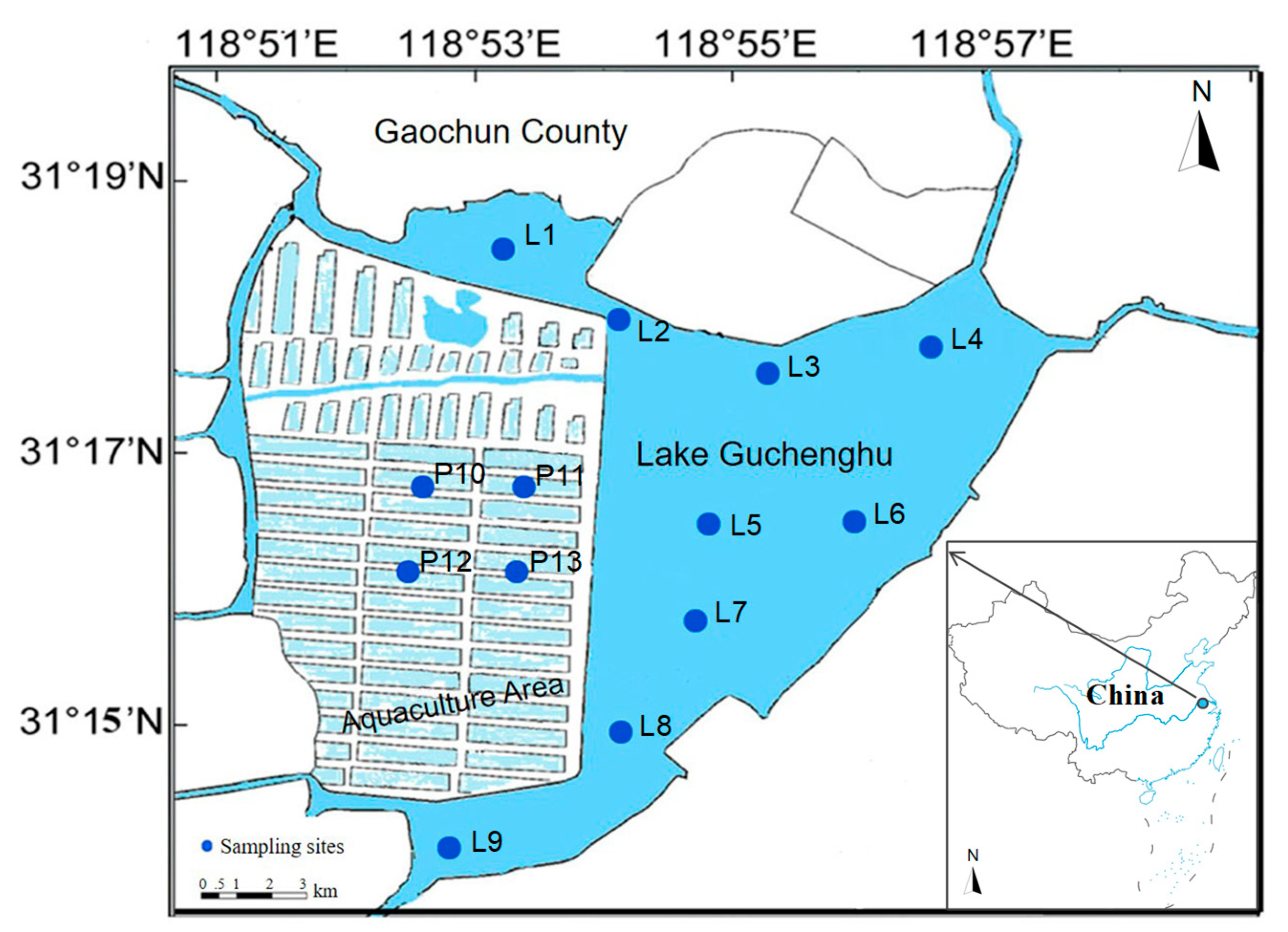
2.1. Study Area
2.2. Sample Collection
2.3. Analysis Methods
2.4. Statistical Analysis
3. Results
3.1. Environmental Parameters
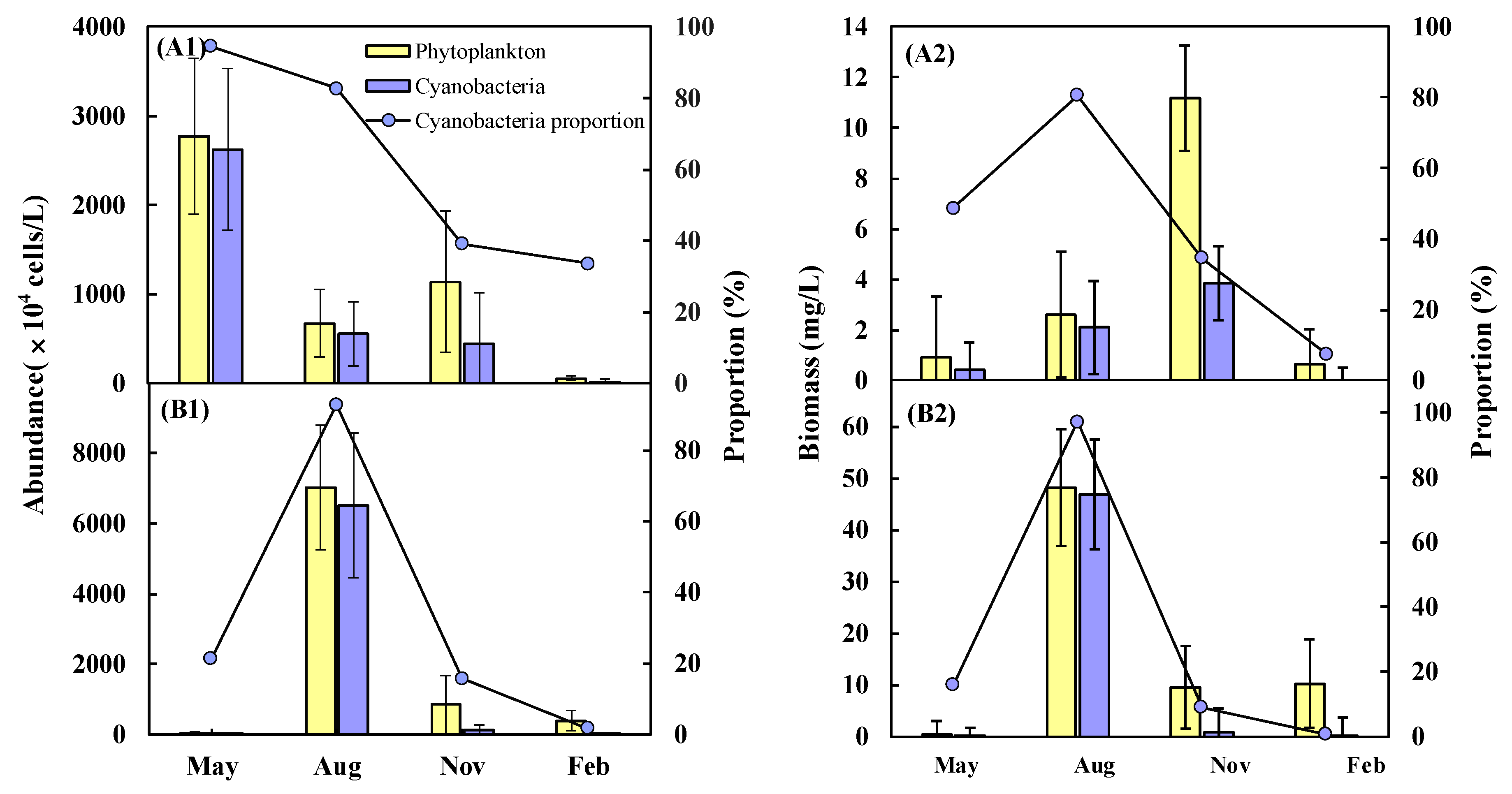
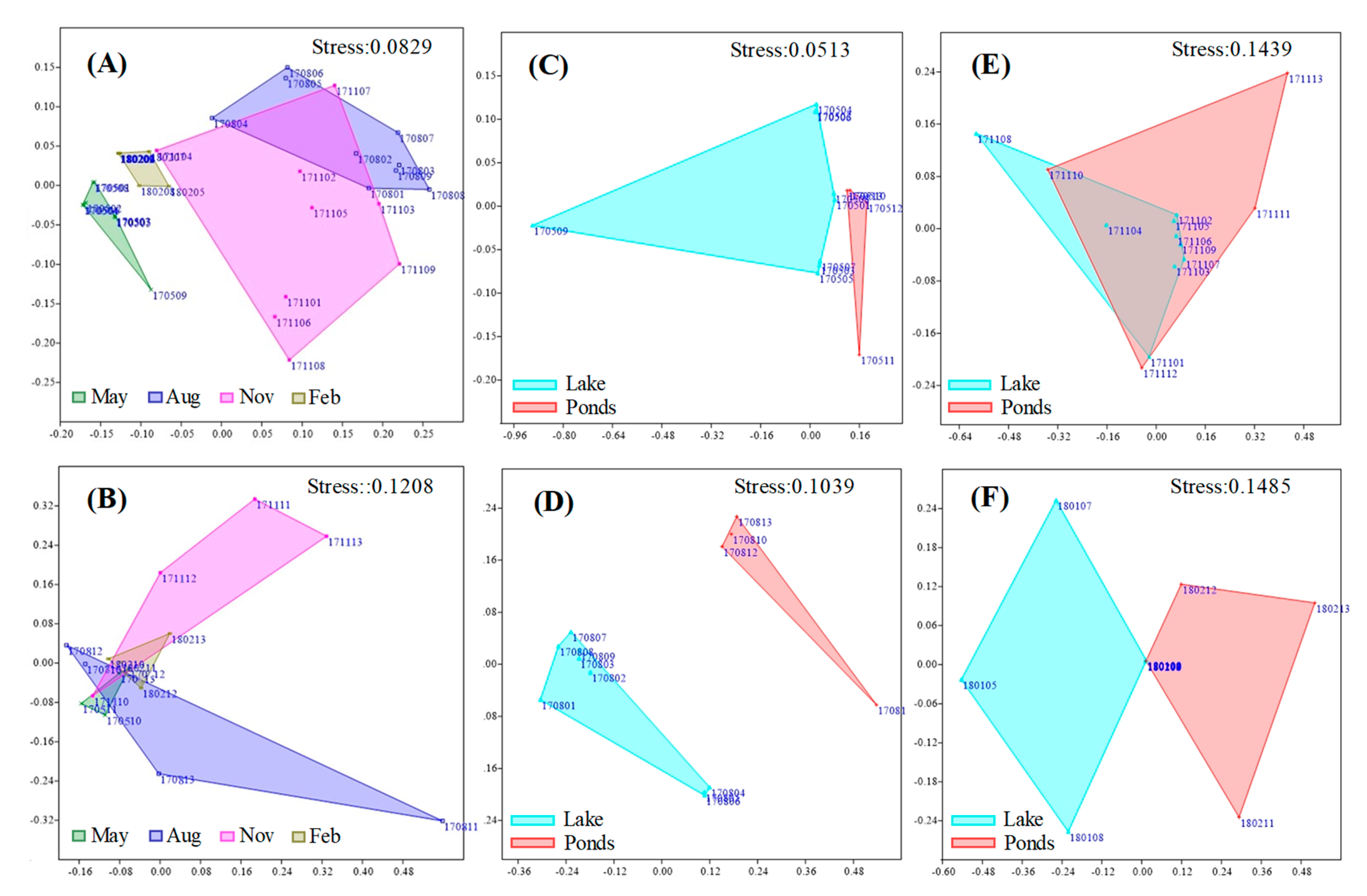
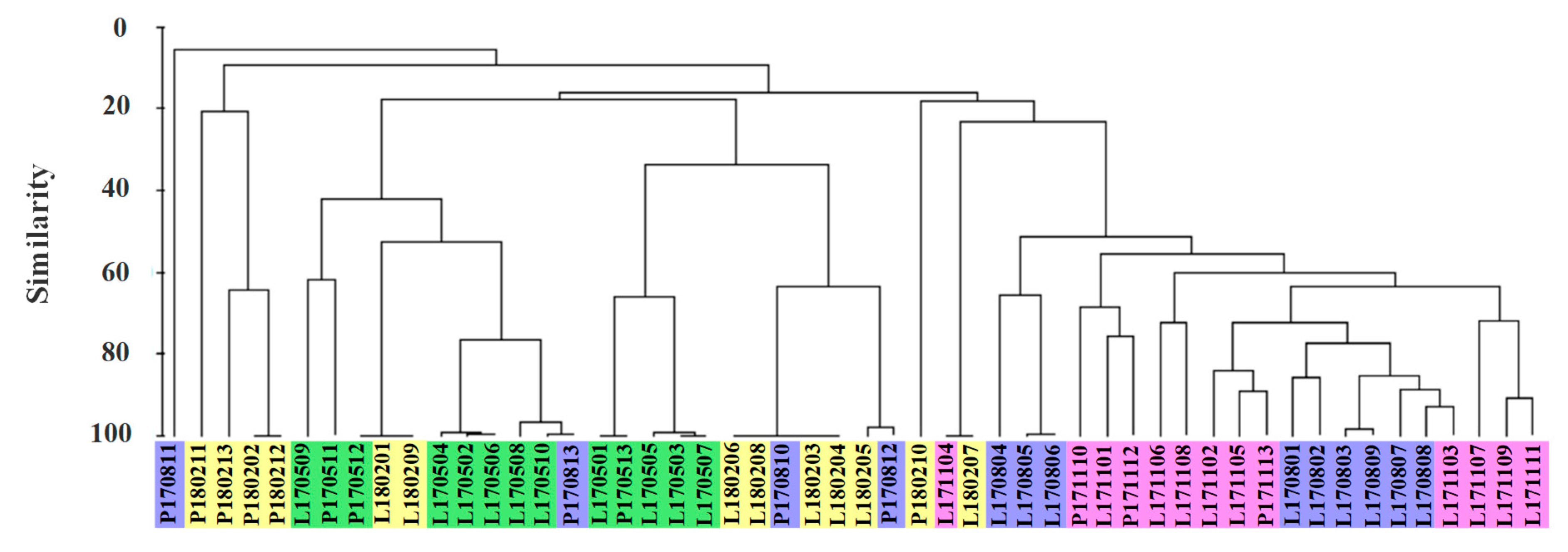
3.2. Seasonal Variations of Cyanobacteria
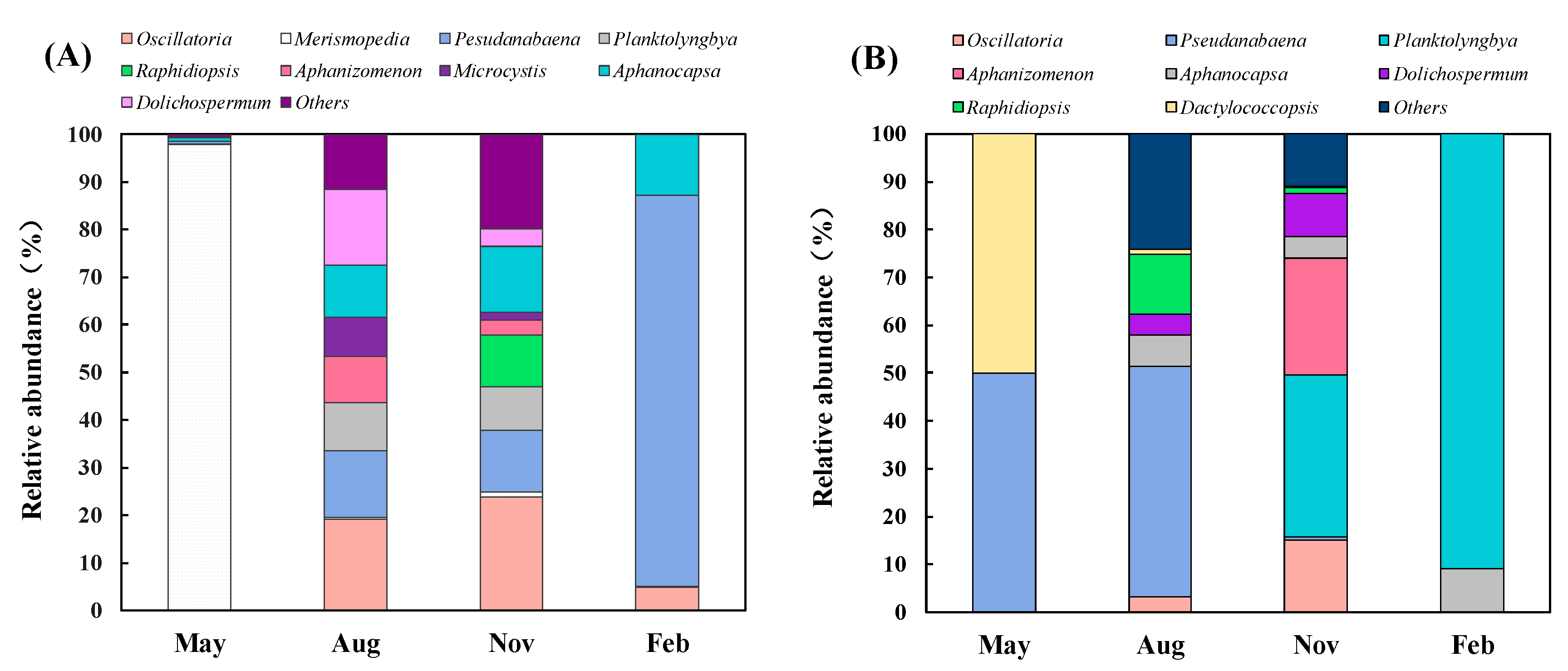
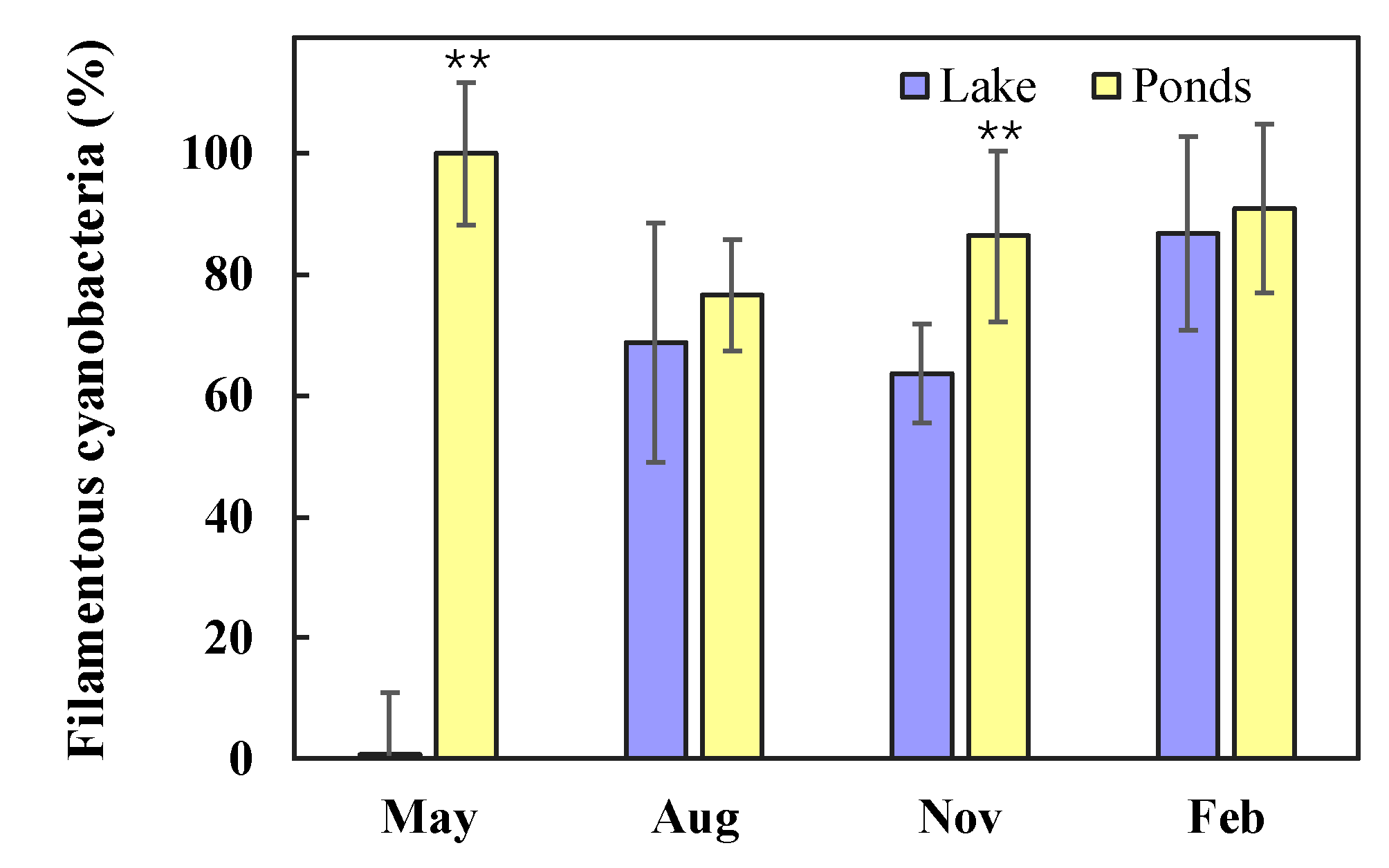
3.3. Quantitative Seasonal Changes in Cyanobacteria Communities
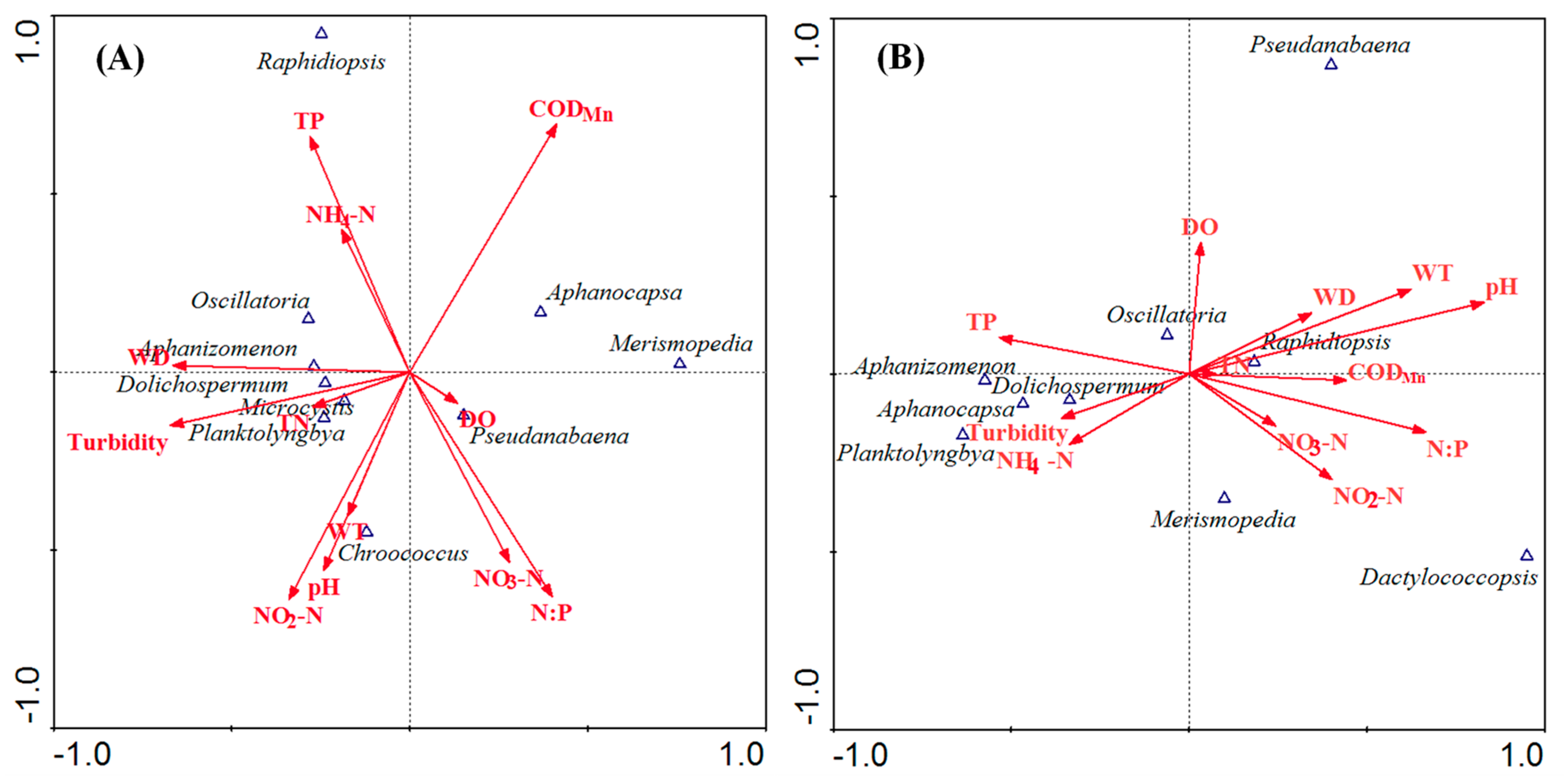
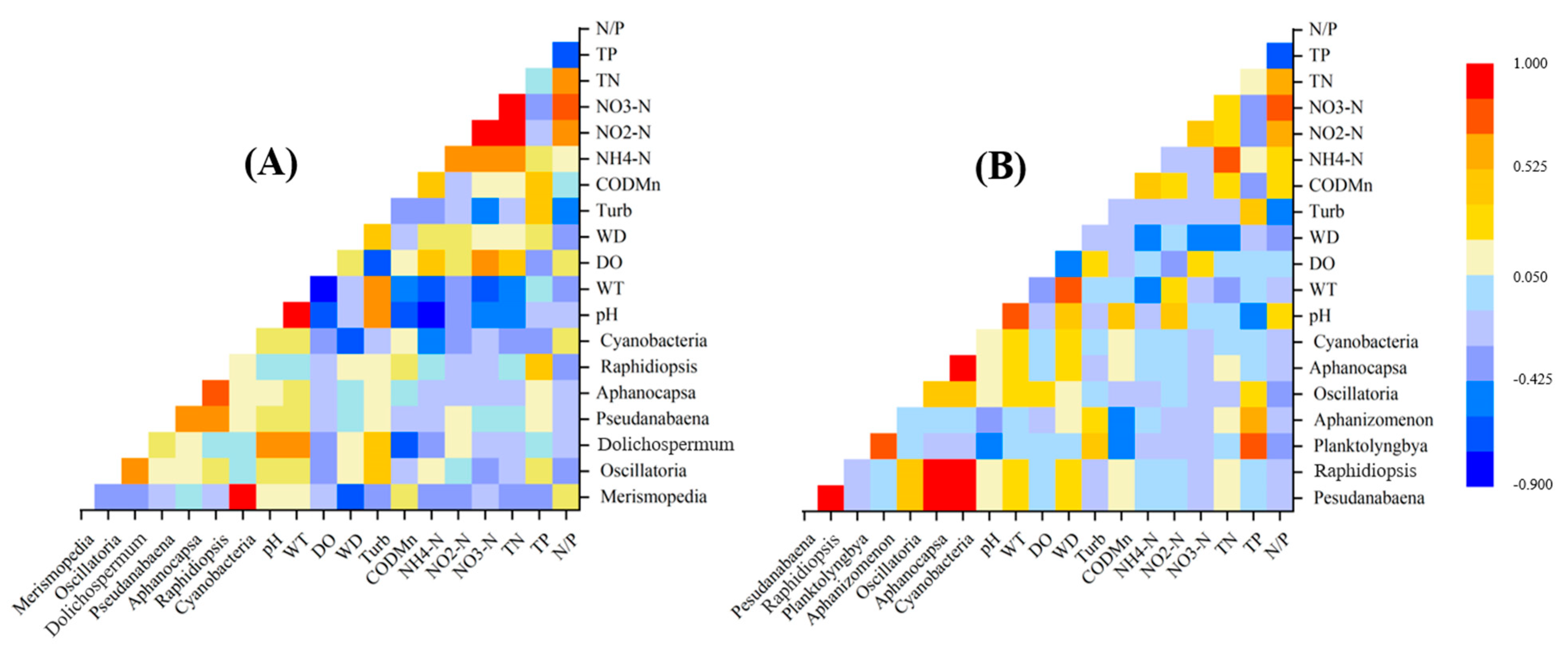
3.4. Relationships between Cyanobacteria Dynamics and Environmental Variables
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cremona, F.; Tuvikene, L.; Haberman, J.; Nõges, P.; Nõges, T. Factors controlling the three-decade long rise in cyanobacteria biomass in a eutrophic shallow lake. Sci. Total Environ. 2018, 621, 352–359. [Google Scholar] [CrossRef]
- Havens, K.E.; Ji, G.; Beaver, J.R.; Fulton, R.S.; Teacher, C.E. Dynamics of cyanobacteria blooms are linked to the hydrology of shallow Florida lakes and provide insight into possible impacts of climate change. Hydrobiologia 2019, 829, 43–59. [Google Scholar] [CrossRef]
- Lenard, T.; Ejankowski, W.; Poniewozik, M. Responses of Phytoplankton Communities in Selected Eutrophic Lakes to Variable Weather Conditions. Water 2019, 11, 1207. [Google Scholar] [CrossRef]
- Hu, X.; Zhang, R.; Ye, J.; Wu, X.; Zhang, Y.; Wu, C. Monitoring and research of microcystins and environmental factors in a typical artificial freshwater aquaculture pond. Environ. Sci. Pollut. Res. 2018, 25, 5921–5933. [Google Scholar] [CrossRef] [PubMed]
- Ndlela, L.L.; Oberholster, P.J.; Van, W.J.H.; Cheng, P. An overview of cyanobacterial bloom occurrences and research in Africa over the last decade. Harmful Algae 2016, 60, 11–26. [Google Scholar] [CrossRef]
- Liao, J.; Zhao, L.; Cao, X.; Sun, J.; Gao, Z.; Wang, J.; Jiang, D.; Fan, H.; Huang, Y. Cyanobacteria in lakes on Yungui Plateau, China are assembled via niche processes driven by water physicochemical property, lake morphology and watershed land-use. Sci. Rep. 2016, 6, 36357. [Google Scholar] [CrossRef]
- Alvarez, S.; Lupi, F.; Solís, D.; Thomas, M. Valuing Provision Scenarios of Coastal Ecosystem Services: The Case of Boat Ramp Closures Due to Harmful Algae Blooms in Florida. Water 2019, 11, 1250. [Google Scholar] [CrossRef]
- Zeng, Q.; Jeppesen, E.; Gu, X.; Mao, Z.; Chen, H. Cannibalism and Habitat Selection of Cultured Chinese Mitten Crab: Effects of Submerged Aquatic Vegetation with Different Nutritional and Refuge Values. Water 2018, 10, 1542. [Google Scholar] [CrossRef]
- Cai, C.; Gu, X.; Huang, H.; Dai, X.; Ye, Y.; Shi, C. Water quality, nutrient budget, and pollutant loads in Chinese mitten crab (Eriocheir sinensis) farms around East Taihu Lake. Chin. J. Oceanol. Limn. 2012, 30, 29–36. [Google Scholar] [CrossRef]
- Zeng, Q.; Gu, X.; Chen, X.; Mao, Z. The impact of Chinese mitten crab culture on water quality, sediment and the pelagic and macrobenthic community in the reclamation area of Guchenghu Lake. Fish. Sci. 2013, 79, 689–697. [Google Scholar] [CrossRef]
- Gu, X.K.; Gu, X.H.; Zeng, Q.F.; Mao, Z.G.; Li, X.G.; Wang, Y.P.; Wang, W.X. Spatial-Temporal Variation and Developing Tendency of Water Quality in Gucheng Lake and Inlets and Outlets of the Lake. J. Ecol. Rural Environ. 2016, 32, 68–75. [Google Scholar]
- Fortin, N.; Aranda-Rodriguez, R.; Jing, H.; Pick, F.; Bird, D.; Greer, C.W. Detection of microcystin-producing cyanobacteria in Missisquoi Bay, Quebec, Canada, using quantitative PCR. Appl. Environ. Microbiol. 2010, 76, 5105–5112. [Google Scholar] [CrossRef] [PubMed]
- Brasil, J.; Attayde, J.L.; Vasconcelos, F.R.; Dantas, D.D.; Huszar, V.L. Drought-induced water-level reduction favors cyanobacteria blooms in tropical shallow lakes. Hydrobiologia 2016, 770, 145–164. [Google Scholar] [CrossRef]
- Yang, C.; Nan, J.; Li, J. Driving factors and dynamics of phytoplankton community and functional groups in an estuary reservoir in the Yangtze River, China. Water 2019, 11, 1184. [Google Scholar] [CrossRef]
- Bullerjahn, G.S.; McKay, R.M.; Davis, T.W.; Baker, D.B.; Boyer, G.L.; Anglada, L.V.; Doucette, G.J.; Ho, J.C.; Irwin, E.G.; Kling, C.L. Global solutions to regional problems: Collecting global expertise to address the problem of harmful cyanobacterial blooms. A Lake Erie case study. Harmful Algae 2016, 54, 223–238. [Google Scholar] [CrossRef]
- Funari, E.; Manganelli, M.; Buratti, F.M.; Testai, E. Cyanobacteria blooms in water: Italian guidelines to assess and manage the risk associated to bathing and recreational activities. Sci. Total Environ. 2017, 598, 867–880. [Google Scholar] [CrossRef]
- Conley, D.J.; Paerl, H.W.; Howarth, R.W.; Boesch, D.F.; Seitzinger, S.P.; Havens, K.E.; Lancelot, C.; Likens, G.E. Controlling eutrophication: Nitrogen and phosphorus. Science 2009, 323, 1014–1015. [Google Scholar] [CrossRef]
- Heisler, J.; Glibert, P.M.; Burkholder, J.M.; Anderson, D.M.; Cochlan, W.; Dennison, W.C.; Dortch, Q.; Gobler, C.J.; Heil, C.A.; Humphries, E. Eutrophication and harmful algal blooms: A scientific consensus. Harmful Algae 2008, 8, 3–13. [Google Scholar] [CrossRef]
- Chan, F.; Pace, M.L.; Howarth, R.W.; Marino, R.M. Bloom formation in heterocystic nitrogen-fixing cyanobacteria: The dependence on colony size and zooplankton grazing. Limnol. Oceanogr. 2004, 49, 2171–2178. [Google Scholar] [CrossRef]
- Lee, T.A.; Rollwagen-Bollens, G.; Bollens, S.M.; Faber-Hammond, J.J. Environmental influence on cyanobacteria abundance and microcystin toxin production in a shallow temperate lake. Ecotoxicol. Environ. Saf. 2015, 114, 318–325. [Google Scholar] [CrossRef]
- Woodhouse, J.N.; Kinsela, A.S.; Collins, R.N.; Bowling, L.C.; Honeyman, G.L.; Holliday, J.K.; Neilan, B.A. Microbial communities reflect temporal changes in cyanobacterial composition in a shallow ephemeral freshwater lake. ISME J. 2016, 10, 1337–1351. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Gu, X.; Mao, Z.; Sun, M.; Gu, X. Assessment of trophic levels and phytoplankton variation in Guchenghu Lake and canal route. Chin. Environ. Sci. 2012, 32, 1487–1494. [Google Scholar]
- Jin, X.C.; Tu, Q.Y. The Standard Methods for Observation and Analysis in Lake Eutrophication; China Environmental Science Press: Beijing, China, 1990; p. 240. [Google Scholar]
- Hu, H. The Freshwater Algae of China: Systematics, Taxonomy and Ecology; Science Press: Beijing, China, 2006. [Google Scholar]
- Hillebrand, H.; Dürselen, C.; Kirschtel, D.; Pollingher, U.; Zohary, T. Biovolume calculation for pelagic and benthic microalgae. J. Phycol. 1999, 35, 403–424. [Google Scholar] [CrossRef]
- Carmichael, W.W. Health Effects of Toxin-Producing Cyanobacteria: “The CyanoHABs”. Hum. Ecol. Risk Assess. Int. J. 2001, 7, 1393–1407. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Walls, J.T.; Wyatt, K.H.; Doll, J.C.; Rubenstein, E.M.; Rober, A.R. Hot and toxic: Temperature regulates microcystin release from cyanobacteria. Sci. Total Environ. 2018, 610, 786–795. [Google Scholar] [CrossRef]
- Pilkaitytė, R.; Razinkovas, A. Seasonal changes in phytoplankton composition and nutrient limitation in a shallow Baltic lagoon. Boreal Environ. Res. 2007, 12, 551–559. [Google Scholar]
- Dalu, T.; Wasserman, R.J. Cyanobacteria dynamics in a small tropical reservoir: Understanding spatio-temporal variability and influence of environmental variables. Sci. Total Environ. 2018, 643, 835–841. [Google Scholar] [CrossRef]
- Qin, X.B.; Huang, P.Y.; Liu, M.H.; Ma, C.X.; Yu, H.X. Phytoplankton assemblage classification in Anbang river wetland in summer. South Chin. Fish. Sci. 2007, 3, 1–7. [Google Scholar]
- Yang, J.; Lv, H.; Liu, L.; Yu, X.; Chen, H. Decline in water level boosts cyanobacteria dominance in subtropical reservoirs. Sci. Total Environ. 2016, 557, 445–452. [Google Scholar] [CrossRef]
- Liu, L.; Liu, D.F.; Johnson, D.M.; Yi, Z.Q.; Huang, Y.L. Effects of vertical mixing on phytoplankton blooms in Xiangxi Bay of Three Gorges Reservoir: Implications for management. Water Res. 2012, 46, 2121–2130. [Google Scholar] [CrossRef] [PubMed]
- Wantzen, K.M.; Rothhaupt, K.O.; Mörtl, M.; Cantonati, M.; László, G.; Fischer, P. Ecological effects of water-level fluctuations in lakes: An urgent issue. Hydrobiologia 2008, 613, 1–4. [Google Scholar] [CrossRef]
- Bothe, H.; Schmitz, O.; Yates, M.G. Newton WE Nitrogen fixation and hydrogen metabolism in cyanobacteria. Microbiol. Mol. Biol. Rev. 2010, 74, 529–551. [Google Scholar] [CrossRef]
- Carey, C.C.; Ibelings, B.W.; Hoffmann, E.P.; Hamilton, D.P.; Brookes, J.D. Eco-physiological adaptations that favour freshwater cyanobacteria in a changing climate. Water Res. 2012, 46, 1394–1407. [Google Scholar] [CrossRef] [PubMed]
- An, W.C.; Li, X.M. Phosphate adsorption characteristics at the sediment—Water interface and phosphorus fractions in Nansi Lake, China, and its main inflow rivers. Environ. Monit. Assess. 2009, 148, 173–184. [Google Scholar] [CrossRef]
- Andersson, A.; Höglander, H.; Karlsson, C.; Huseby, S. Key role of phosphorus and nitrogen in regulating cyanobacterial community composition in the northern Baltic Sea. Estuar. Coast. Shelf Sci. 2015, 164, 161–171. [Google Scholar] [CrossRef] [Green Version]
- Paerl, H.W.; Paul, V.J. Climate change: Links to global expansion of harmful cyanobacteria. Water Res. 2012, 46, 1349–1363. [Google Scholar] [CrossRef]
- Dokulil, M.T.; Teubner, K. Cyanobacterial dominance in lakes. Hydrobiologia 2000, 438, 1–12. [Google Scholar] [CrossRef]
- Antunes, J.T.; Leão, P.N.; Vasconcelos, V.M. Cylindrospermopsis raciborskii: Review of the distribution, phylogeography, and ecophysiology of a global invasive species. Front. Microbiol. 2015, 6, 473. [Google Scholar] [CrossRef] [Green Version]
- Kozak, A.; Celewicz-Gołdyn, S.; Kuczyńska-Kippen, N. Cyanobacteria in small water bodies: The effect of habitat and catchment area conditions. Sci. Total Environ. 2019, 646, 1578–1587. [Google Scholar] [CrossRef]
- Drobac, D.; Tokodi, N.; Lujić, J.; Marinović, Z.; Subakov-Simić, G.; Dulić, T.; Važić, T.; Nybom, S.; Meriluoto, J.; Codd, G.A. Cyanobacteria and cyanotoxins in fishponds and their effects on fish tissue. Harmful Algae 2016, 55, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Peng, L.; Huang, X.; Han, B. Occurrence and dominance of Cylindrospermopsis raciborskii and dissolved cylindrospermopsin in urban reservoirs used for drinking water supply. South China Environ. Monit. Assess. 2014, 186, 3079–3090. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Chen, H.; Liu, M.; Yang, J.R.; Xiao, P.; Wilkinson, D.M.; Yang, J. Response of the eukaryotic plankton community to the cyanobacterial biomass cycle over 6 years in two subtropical reservoirs. ISME J. 2019, 13, 2196–2208. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | May, 2017 | August, 2017 | November, 2017 | February, 2018 | ||||
|---|---|---|---|---|---|---|---|---|
| Lake | Ponds | Lake | Ponds | Lake | Ponds | Lake | Ponds | |
| pH | 8.23 ± 0.19 | 8.76 ± 0.42 * | 9.15 ± 0.17 | 8.24 ± 0.41 * | 7.61 ± 0.23 | 7.06 ± 0.25 * | 7.43 ± 0.13 | 7.45 ± 0.06 |
| WT/(°C) | 21.15 ± 0.75 | 24.93 ± 1.81 * | 29.47 ± 0.31 | 31.01 ± 1.54 * | 18.37 ± 0.25 | 18.40 ± 0.18 | 3.10 ± 0.14 | 3.12 ± 1.53 |
| DO/(mg/L) | 12.43 ± 2.04 | 10.23 ± 0.75 * | 11.19 ± 0.39 | 16.49 ± 4.82 * | 10.37 ± 0.76 | 12.89 ± 4.75 | 13.68 ± 7.33 | 16.90 ± 2.67 * |
| WD/(m) | 2.82 ± 0.89 | 1.11 ± 0.02 * | 4.12 ± 0.55 | 1.13 ± 0.01 * | 4.19 ± 0.33 | 0.98 ± 0.02 * | 4.05 ± 0.31 | 0.74 ± 0.27 * |
| Turb/(NTU) | 7.02 ± 4.84 | 19.5 ± 7.81 * | 26.3 ± 12.08 | 27.9 ± 6.93 | 18.9 ± 1.62 | 45.2 ± 24.8 * | 4.46 ± 1.10 | 45.9 ± 19.71 * |
| NO2−-N/(mg/L) | 0.03 ± 0.00 | 0.02 ± 0.01 * | 0.04 ± 0.01 | 0.025 ± 0.01 | 0.02 ± 0.01 | 0.01 ± 0.01 | 0.07 ± 0.05 | 0.03 ± 0.03 |
| −-N/(mg/L) | 0.56 ± 0.07 | 0.32 ± 0.28 * | 0.45 ± 0.14 | 0.23 ± 0.18 * | 0.13 ± 0.04 | 0.16 ± 0.13 | 1.87 ± 1.11 | 0.67 ± 1.01 |
| NH4+-N/(mg/L) | 0.05 ± 0.02 | 0.34 ± 0.30 * | 0.05 ± 0.02 | 0.17 ± 0.14 * | 0.11 ± 0.04 | 0.17 ± 0.14 | 0.15 ± 0.03 | 2.51 ± 3.81 |
| TN/(mg/L) | 1.02 ± 0.11 | 2.00 ± 0.54 * | 1.18 ± 0.17 | 1.57 ± 0.48 * | 1.25 ± 0.07 | 1.66 ± 0.75 | 2.47 ± 1.49 | 3.04 ± 2.62 |
| TP/(mg/L) | 0.02 ± 0.01 | 0.09 ± 0.02 * | 0.03 ± 0.01 | 0.18 ± 0.10 * | 0.22 ± 0.02 | 0.36 ± 0.11 * | 0.04 ± 0.00 | 0.14 ± 0.08 * |
| N:P | 59.6 ± 17.6 | 22.5 ± 9.62 * | 35.5 ± 4.74 | 11.4 ± 5.75 * | 5.75 ± 0.34 | 4.39 ± 0.87 * | 62.5 ± 41.8 | 23.0 ± 16.8 |
| CODMn/(mg/L) | 4.14 ± 0.30 | 9.19 ± 1.31 * | 2.86 ± 0.19 | 6.22 ± 1.28 * | 4.37 ± 0.18 | 4.64 ± 2.07 | 4.05 ± 0.61 | 7.62 ± 3.03 |
| Species a | Family | Maximum Abundance | |
|---|---|---|---|
| Lake | Ponds | ||
| Planktothrix agardhii | Oscillatoriaceae | 1.1 × 105 cells/L in November | -- |
| Dolichospermum circinale (synonym: Anabaena circinale) | Nostocaceae | 1.2 × 105 cells/L in August | 3.6 × 106 cells/L in August |
| Raphidiopsis raciborskii(synonym: Cylindrospermopsis raciborskii) | Nostocaceae | -- | 8.2 × 106 cells/L in August |
| Cuspidothrix issatschenkoi (synonym: Aphanizomenon issatschenkoi”) | Nostocaceae | -- | 4.5×104 cells/L in November |
| Aphanizomenon flos-aquae | Nostocaceae | 1.9 × 105 cells/L in August | -- |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Chen, H.; Gu, X.; Mao, Z.; Zeng, Q.; Ding, H. Dynamics of Cyanobacteria and Related Environmental Drivers in Freshwater Bodies Affected by Mitten Crab Culturing: A Study of Lake Guchenghu, China. Water 2019, 11, 2468. https://doi.org/10.3390/w11122468
Li H, Chen H, Gu X, Mao Z, Zeng Q, Ding H. Dynamics of Cyanobacteria and Related Environmental Drivers in Freshwater Bodies Affected by Mitten Crab Culturing: A Study of Lake Guchenghu, China. Water. 2019; 11(12):2468. https://doi.org/10.3390/w11122468
Chicago/Turabian StyleLi, Hongmin, Huihui Chen, Xiaohong Gu, Zhigang Mao, Qingfei Zeng, and Huiping Ding. 2019. "Dynamics of Cyanobacteria and Related Environmental Drivers in Freshwater Bodies Affected by Mitten Crab Culturing: A Study of Lake Guchenghu, China" Water 11, no. 12: 2468. https://doi.org/10.3390/w11122468