The Toxicity and Uptake of As, Cr and Zn in a Stygobitic Syncarid (Syncarida: Bathynellidae)


1
Department of Biological Sciences, Macquarie University, Sydney, NSW 2109, Australia
2
Department of Molecular Sciences, Macquarie University, Sydney, NSW 2109, Australia
3
Australian Nuclear Science and Technology Organisation, Locked Bag 2001, Kirrawee DC, NSW 2232, Australia
*
Author to whom correspondence should be addressed.
Water 2019, 11(12), 2508; https://doi.org/10.3390/w11122508
Submission received: 22 October 2019
/
Revised: 15 November 2019
/
Accepted: 21 November 2019
/
Published: 28 November 2019
(This article belongs to the Special Issue Assessment of Different Contaminants in Freshwater: Origin, Fate, and Ecological Impact)
Abstract
:Ecotoxicological data for obligate groundwater species are increasingly required to inform environmental protection for groundwater ecosystems. Bathynellid syncarids are one of several crustacean taxa found only in subsurface habitats. The aim of this paper is to assess the sensitivity of an undescribed syncarid (Malacostraca: Syncarida: Bathynellidae) to common groundwater contaminants, arsenic(III), chromium(VI) and zinc, and examine the bioaccumulation of As and Zn in these animals after 14-day exposure. Arsenic was the most toxic to the syncarid (14-day LC50 0.25 mg As/L), followed closely by chromium (14-day LC50 0.51 mg Cr/L) and zinc (14-day LC50 1.77 mg Zn/L). The accumulation of Zn was regulated at exposure concentrations below 1 mg Zn/L above which body concentrations increased, leading to increased mortality. Arsenic was not regulated and was accumulated by the syncarids at all concentrations above the control. These are the first published toxicity data for syncarids and show them to be among the most sensitive of stygobitic crustaceans so far tested, partly due to the low hardness of the groundwater from the aquifer they inhabit and in which they were tested. The ecological significance of the toxicant accumulation and mortality may be significant given the consequent population effects and low capacity for stygobitic populations to recover.
1. Introduction
Groundwater is vital to society globally. It provides drinking water to over 2 billion people worldwide [1], and, in many areas, is the only reliable water source. Increasing demand for water and risk of contamination from surface activities such as agriculture, urban development and mining threaten the integrity of aquifers, their suitability as a water source, and the biota that inhabit these subterranean ecosystems [2].
Groundwater ecosystems support a unique biota, consisting of microbial assemblages, frequently crustacean-dominated invertebrate assemblages and, somewhat rarely, vertebrates such as fish [3]. Groundwater invertebrates particularly have evolved to dark, stable and low energy conditions of groundwater environments, such that aquifers the world over contain a diversity of unique taxa not found in surface environments [3]. Bathynellid syncarids are archetypal groundwater organisms. Found only in hyporheic or groundwater environments [4], bathynellid syncarids share the common morphological traits of blindness, lack of body pigments, vermiform (wormlike) body shape, and enhanced sensory appendages that have evolved independently across multiple lineages of groundwater fauna [3]. These traits and other adaptations make groundwater fauna morphologically and physiologically different from even related surface water species, and consequently, they may be expected to respond differently to toxicants compared to surface-dwelling species [5].
As in any ecosystem, pollution threatens the health of the groundwater biota, with flow-on effects to other elements of the ecosystem. For groundwater ecosystems, disruption to one element of the ecosystem is likely to have significant effects on the microbial communities and subsequently impact the ability of the ecosystem to self-purify and provide clean groundwater [6]. The risk to groundwater ecosystems is that the fauna has evolved classic k-strategist traits of longevity, low metabolic and reproductive rates [3], meaning that recovery from disturbance is, at best, slow [7].
Contamination of groundwater by metals is a global concern, often occurring as a result of industrial uses, spills and land application of contaminated materials (Castano-Sanchez in press) [8]. Unfortunately, groundwater ecosystems may not be protected by water quality legislation to the same extent as other aquatic ecosystems (such as rivers and estuaries) because water quality guidelines for ecosystem protection are not based on toxicological data for the biota of that ecosystem [9,10]. Instead, existing water quality guidelines for groundwater ecosystems are based on toxicity data for surface waters, but there are a number of reasons why groundwater biota may respond differently to toxicants compared to surface water taxa [5]. In particular, the metabolic rates of groundwater macroinvertebrates tend to be lower than those of related epigean species [11]. As a result, the uptake of toxicants may be lower where uptake of toxicants is an active, metabolic-driven process [5]. Alternatively, the removal of toxicants accumulated by passive processes may be low, leading to greater body burdens in groundwater fauna compared to surface water fauna [5].
The aim of this study is to test the sensitivity of an obligate groundwater crustacean (Malacostraca: Syncarida) to metal and metalloid contaminants (Zn, As and Cr) in groundwater. The first step to mortality and harm from contaminants is the uptake of those contaminants by the organism, so we also examine here the uptake of these metals by the syncarids and relate uptake to exposure and mortality.
2. Materials and Methods
2.1. Test Species
In this study, we test an undescribed species of the family Bathynellidae (Malacostraca: Syncarida). Until recently, all Australian Bathynellidae were assigned to the genus Bathynella, but phylogenies using newly collected specimens suggest strong regional separation within the Australian Bathynellidae, with specimens from eastern Australia being a separate clade [12,13,14], and those tested here likely a novel taxon. Type specimens are held at Macquarie University. Unfortunately, taxonomic keys to allow further identification are currently unavailable. However, multiple specimens from each test were examined to ensure morphological consistency of the test organisms within and between tests. Syncarida do not have morphologically different life stages after hatching [4], making it difficult to determine life stage; however, the syncarids tested were approximately 1000 µm long and of similar size within and between tests.
Test animals were collected from a fractured sandstone aquifer at Somersby, NSW Australia, approximately 80 km north of Sydney (33°22′15.4″ S, 151°18′9″ E, Bore depth = 22 m). The sample collection area is semirural. Groundwater in the region is used heavily by beverage bottling industries and is considered of very high quality. There is no recorded history of groundwater contamination at the site and analysis of the water over time has revealed very low concentrations of As, Cr and Zn, as seen in Table 1. The groundwater is typically low in dissolved solids and has a slightly acidic pH as a consequence of iron in the sandstone aquifer, as seen in Table 1.
Syncarids were collected from groundwater as described in Korbel & Hose [15] as summarised below. Groundwater was pumped using a motorised inertia pump (Waterra, ON, Canada). Approximately 300 L of water was pumped and passed through a 63 µm mesh sieve to collect the invertebrates. The sieve contents were placed in a sealable, 1 L plastic container which was filled with clean groundwater and placed in a portable cooler for transportation to the laboratory.
In the laboratory, containers containing the invertebrates were placed in a dark environmental cabinet at 18 °C, which approximates the temperature of the groundwater at the time of collection. Syncarids were acclimated to conditions in the environmental cabinet for at least 48 h prior to testing. Tests were conducted within seven days of collection and data from repeat tests were pooled for analysis.
Tests were conducted following the methods reported in Hose et al. [16], which are summarised below. Once acclimated, the syncarids were removed from the collection container using a pipette and placed individually into wells of 24-well plates (Greiner) along with a known volume of clean groundwater from the collection site. Treatment concentrations were randomly allocated to each well and the required volume of clean groundwater added such that the subsequent addition of toxicant solution would give a final test volume of 2.5 mL. The 24-well plates were covered and placed in a dark cabinet at 18 °C for the duration of the test. Mortality was monitored daily and dead animals were removed, rinsed in Milli-Q water and frozen individually for later analysis.
Toxicity tests were conducted over 14 days with the test end point being immobility, defined as failure to move within 15 s of being gently prodded. This test was repeated twice within 15 min to confirm immobility. Immobility was recorded at 96 h and 14 days. Dissolved oxygen, conductivity, pH and temperature were recorded every 48 h from a randomly selected replicate of each treatment using HANNA (Hanna Instruments Inc, Smithfield, RI, USA) handheld meters.
Each test was comprised of a negative control and six or seven test concentrations, each with four or three replicate wells per plate, respectively. The number of animals varied between tests depending on the number in field collections and controls in all tests had ≥13 individuals. The concentration ranges used for each toxicant were 0–10 mg As/L, 0–100 mg Zn/L, and 0–10 mg Cr/L. These concentrations span the ranges reported in groundwater in the literature (e.g., [17,18,19,20]). Toxicants were tested individually. Results from a test were not accepted if there was more than 20% mortality of organisms in control treatments [10].
Toxicant stock solutions were made up in Milli-Q water. Chromium(VI) was added as K2Cr2O7 (Sigma-Aldrich, 99% purity), arsenic was added as NaAsO2 (Sigma-Aldrich, 98% purity) and zinc was added as ZnSO4.7H2O (Ajax Finechem, 99% purity). Test concentrations were measured using the S2 Picofox Total Reflection X-Ray Fluorescence (TRX-RF) Spectrometer (Bruker Nano GMBH, Berlin, Germany), using the method for aqueous samples as outlined in [21]. A 1 mL aliquot of toxicant solution was taken from randomly selected wells of each treatment at the completion of each test. The aliquot of toxicant solution was spiked with approximately 5–10 µL of standard gallium solution, which acts as an internal standard [21,22]. Aliquots of 20 µL of this solution were then pipetted onto clean, hydrophobic carriers and dried before analysis. Analyses of spectra was done using the Spectra software (version 7.5.3.0, Bruker Nano GMBH, Berlin, Germany). Toxicant concentrations were averaged across replicates and these average values were used in subsequent statistical analyses. Merck XVI multielement standard (Merck KGaA, Darmstadt, Germany) was analysed along with the samples, and indicated average (±SD) recoveries of 107 ± 8% for As, 95 ± 5% for Cr, and 80 ± 5% for Zn. Limits of detection were 1 µg/L for As and Zn, and 3 µg/L for Cr.
The accumulation of As and Zn in the syncarids was measured using Particle Induced X-ray Emission (PIXE). Individual syncarids were freeze dried and placed whole, intact on to sample holders (see below). Resource limitations precluded the analysis of Cr.
2.2. PIXE Elemental Analysis
The samples were analysed for trace and bulk element concentrations with μ-PIXE using the Heavy Ion Microprobe (HIMP) at the Australian Nuclear and Science Technology Organisation (ANSTO) [23]. The HIMP uses the Australian National Tandem Research (ANTARES) accelerator to accelerate ions to the MeV energy range. In this analysis, a 3 MeV proton beam with typical spot size of between 5 and 10 μm was used. At this spot size, beam currents between 0.4 and 0.5 nA can be achieved, which is sufficient for μ-PIXE analyses. A high-purity Germanium (Ge) detector was used with a 100 mm2 active area to measure the characteristic x-rays emerging from the calcification samples. The detector was located 32 mm from the sample for optimum count rate and sensitivity. A 100 μm Mylar foil was used to reduce the low energy x-rays count as well as prevent scattered protons from entering the detector. Samples were fixed to the sample holder by means of double-sided carbon tape. This tape is very pure and does not contain trace elements higher than oxygen (Z > 8). The tape does not affect measurements and is effective in holding the samples providing good electric conductivity to allow target current measurement. Each sample was then scanned for 5–30 min for extensive emission data to be obtained. Elemental concentration maps were created using GeoPIXE [24,25] and used to calculate the average elemental concentrations.
2.3. Statistical Analysis
Measured toxicant concentrations were compared with nominal concentrations using least squares linear regression. Concentration response curves were estimated by fitting a two-parameter nonlinear regression function with a binomial error structure using the DRC package [26] in R version 3.5.3 [27]. Weibull, log-logistic and log-normal models were fit to each dataset, with the best fitting model selected based on Akaikes Information Criteria. The model parameters were estimated using maximum likelihood, with starter values determined by the programs self-starter function. Concentrations affecting 10% and 50% of the syncarids (EC10 and EC50 values, respectively) were extrapolated from the fitted curve. Data from both 96 h and 14-day exposures were analysed separately.
The concentrations of As and Zn measured in syncarids after exposure were compared between treatments using one-way analyses of variance. Post-hoc comparisons between treatments were made using SNK tests. Data were log transformed to approximate normality and homogeneity of variance, which were assessed visually using plots of residuals and Q-Q plots. Analyses of variance were done using the NCSS program (Version 10, NCSS LLC, Kaysville, Utah, USA). The significance level (α) for all analyses was 0.05.
3. Results
Measured test concentrations were mostly within 20% of the nominal concentrations, and have been used in all analyses. Dissolved oxygen concentrations remained at 60% saturation or higher throughout all tests. This level of oxygen saturation is in line with the general concentrations observed in the field, as seen in Table 1. Temperature was consistently at or close to the test temperature of 18 °C. The hardness of the diluent water was relatively low, as seen in Table 1, which reflects the low carbonate concentrations in the sandstone aquifer from which the fauna and diluent water was collected. pH values varied little across the tests, and were in the range 5–6 pH units, as expected given the naturally acidic groundwater used as the diluent water.
Control mortality at 96 h was below zero for Cr and As exposures, and 3% for Zn. At 14 days, control mortality was higher across all tests (As: 9%, Cr: 11%, Zn: 11%), but below acceptability criterion of 20%, as seen in Figure S1. A Weibull model was fitted to all data sets, with details provided in Figure S1 and Table S1. Arsenic and Cr were similarly toxic after 96 h, but As was the most toxic after 14-day exposure. Zinc was the least toxic to the syncarids across both exposure periods, as seen in Table 2. As expected, toxicity increased and EC values decreased with exposure period from 96 h to 14 days for all toxicants, as seen in Table 2.
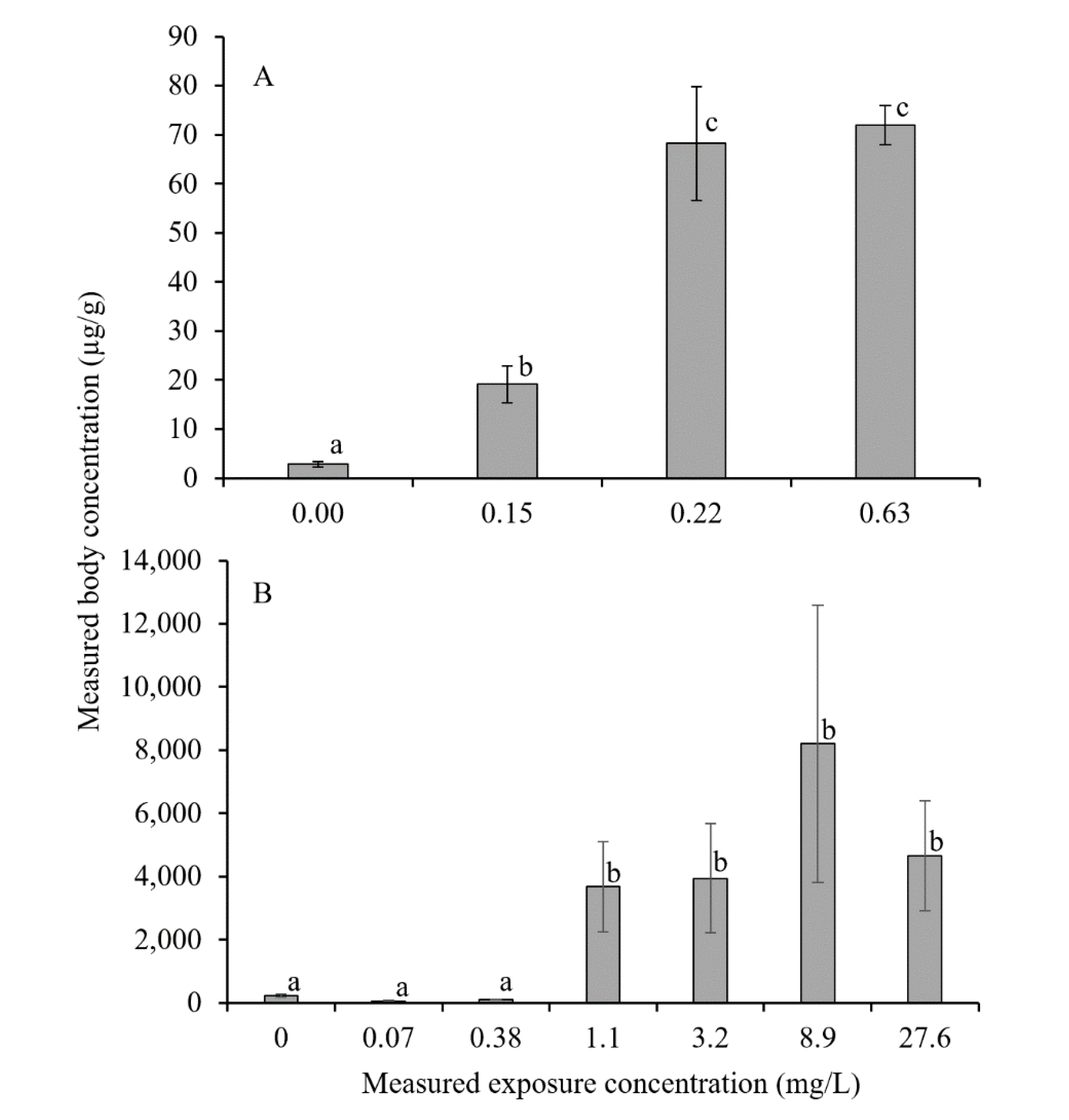
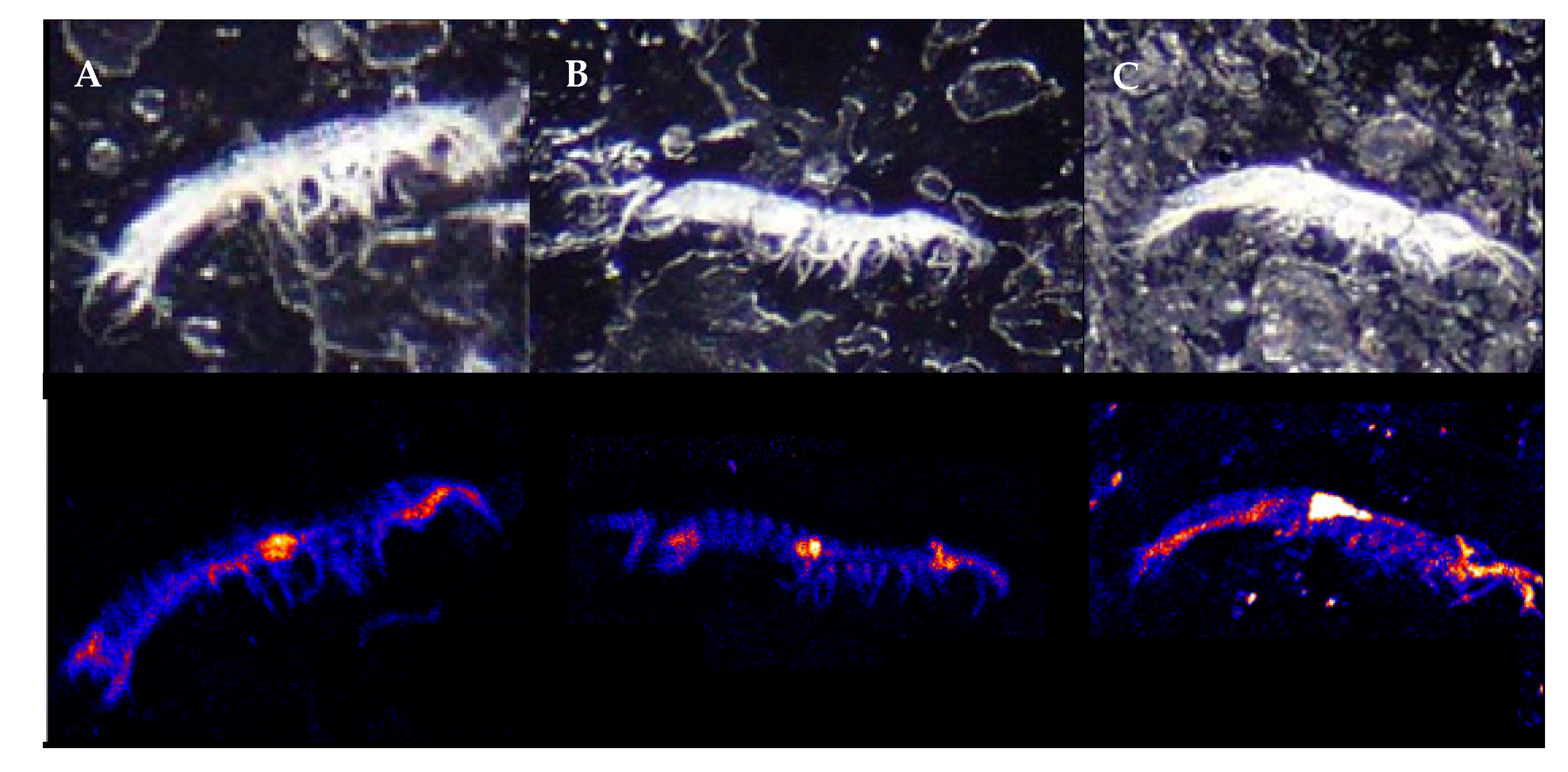
The mean body concentration of As in the syncarids in controls was 2.8 µg As/g, as seen in Figure 1A. The body concentrations in all exposure treatments were significantly greater than in the controls (p < 0.05), and concentrations in syncarids from the 10,000 and 300 mg/L treatments were greater than those in the 100 mg/L treatment (p < 0.05), but were not significantly different to each other, as seen in Figure 1A. The PIXE maps showed that As was concentrated around the head, anterior thoracic and abdominal sections, as seen in Figure 2.
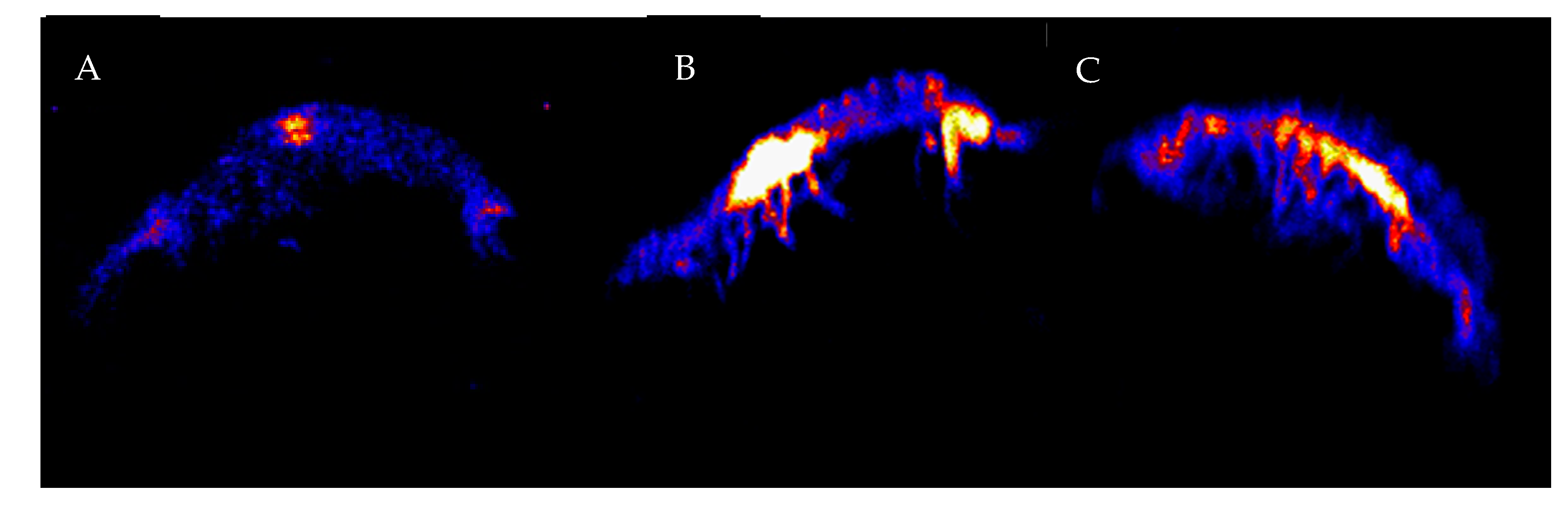
Concentrations of Zn measured in the syncarids from the control treatments were on average 228 µg Zn/g, which increased significantly (p < 0.001) with exposure to all Zn treatments ≥1 mg Zn/L, as seen in Figure 1B. There was no significant difference (p > 0.05) in body concentrations across the Zn treatments, as seen in Figure 1B above 1 mg Zn/L. The accumulation of Zn in the syncarids appears to be concentrated in the anterior thoracic region and abdomen, as seen in Figure 3.
4. Discussion
Our tests have provided the first toxicity and accumulation data for the Syncarida, specifically the Bathynellidae, which are an obligate subterranean taxon. We discuss below the sensitivity of the syncarids to the metals and metalloid relative to other stygobiotic taxa and surface water crustaceans. The accumulation of Zn suggests the ability of syncarids to regulate body concentrations at low exposure concentrations, but for As, we found no evidence of regulation at the exposure concentration tested. Tests were conducted in relatively low hardness, low organic content water from a fractured rock aquifer, which likely makes them more susceptible to toxicants than stygobitic taxa from hard, karstic or more organic groundwaters [28]. While this trend is not unexpected, we highlight the need to consider the aquifer types in the management and risk assessment for subterranean ecosystems.
Mortality in controls was ≤11%, below the acceptable threshold of 20%, and water quality conditions were stable during all tests, indicating we had created a suitable testing environment. Dissolved oxygen (DO) concentrations in the tests were in line with those measured at the collection site in groundwater, as seen in Table 1, but higher than often reported in groundwaters [29]. The relatively high dissolved oxygen concentrations in the static tests is a likely consequence of the small test volume used and large surface area for gas exchange within the 24 well plates. Previous studies suggest that stygobitic crustaceans can survive well in oxygenated laboratory environments (e.g., [11,16]), and do not affect the toxic response of groundwater crustaceans to metals [30].
The toxicity of As to the syncarids after 96 h was similar to that observed for other stygobitic crustaceans. Hose et al. [16] tested the sensitivity of cyclopoid and harpacticoid copepods from the same Somersby location and showed 96-h LC50 values of similar magnitude (5.24, 3.04 mg As/L, respectively) to those estimated here for the syncarids. The syncarids were similar in sensitivity to As as the cladoceran Daphnia magna (96-h LC50 1.50–4.34 mg As(III)/L [31]). However, over 14 days, our syncarids (14-day LC50, 0.25 mg As/L) were slightly more sensitive than the cyclopoid and harpacticoid copepods from Somersby (14-day LC50 0.79, 1.46 mg As/L, respectively [16]). The syncarids were also more sensitive to As than the stygobitic amphipod Niphargus rhenorhodanensis (240-h LC50, 3.97 mg/L [32]), but similar to other freshwater crustaceans such as Gammarus fossarum (LC50, 0.2 mg As/L [32]), which was the most sensitive of a range of taxa in that study. Overall the syncarids were less sensitive to As than Cr, which is consistent with the findings of Canivet et al. [32]. For As and the metals tested, there are few low ECx values in the literature for comparison, nevertheless, LC10 values are reported here so that they may be available to inform the development of environmental quality criteria for groundwaters (e.g., [9]).
Chromium was the most toxic of the elements tested. The acute toxicity of chromium to the syncarids was similar to that recorded for subterranean crustacea reported previously. The ranges of 96-h LC50 values for the styogophilous Bryocamptus echinatus (1.26 mg Cr/L [33]), N. rhenorhodanensis (1.51–2.07 mg Cr/L [32]), stygobitic copepods (0.7–3.1 mg Cr/L [16]), and the stygobitic isopod Proassellus spp. (48-h EC50 0.396–6.35 mg Cr/L [34]) all overlap the 96-h LC50 value for the syncarids (1.8 mg Cr/L). The 96-h LC50 values for D. magna exposed to Cr (0.07–0.16 mg Cr/L, [35,36]) were approximately an order of magnitude lower than those for the syncarids tested here, as seen in Table 2. The 14-day LC50 value (0.5 mg Cr/L) is also within the range of values for copepods from the same study site (0.03–1.06 mg Cr/L [16]) and a 10-day LC50 for N. rhenorhodanensis (0.23 mg Cr/L [32]) and less than those for a range of epigean taxa [32,36]. The 14-day LC10 for Cr (0.004 mg Cr/L) was below the background concentration at the site (0.03 mg Cr/L). This anomaly is a likely consequence of the exposure-response curve fitting, as seen in Figure S1, which is often less reliable at the extremities of the curve. We do, however, believe that the overall curve fit is reliable and the inherent conservatism in this case is preferable for the application of these data to ecosystem protection.
The syncarids were less sensitive to Zn than the Harpacticoid copepod and more sensitive than the cyclopoid copepod from the same locations over both 96 h and 14 days [16]. Ninety-six hour LC50 values for the syncarids were similar to those determined for the groundwater copepod Parastenocaris germanica (96-h LC50 1.7 mg Zn/L) under oxic (10 mg O2/L) conditions [30], but both P. germanica and the syncarid tested here were much more sensitive than several other stygobitic crustaceans such as Niphargus aquilex (96-h LC50 180 mg Zn/L [37]), Asellus cavaticus (96-h LC50 127 mg Zn/L [37]) and Niphargus montellianus, which had no mortality to Zn after 10-day exposure to 25 mg Zn/L [38]. The syncarids were less sensitive to zinc over 96 h than was D. magna (96-h LC50 0.15–0.18 mg Zn/L [36,39]).
Overall, the syncarids tested were similar, but slightly more sensitivity than other stygobitic crustaceans. The syncarids were, however, generally less sensitive over 96 h exposure than the standard cladoceran test species, D. magna. Current approaches for risk assessment for groundwater include the use of standard test species, such as D. magna [40], as a first tier in lieu of toxicity data for groundwater biota. Our results suggest that this may be a suitable interim approach until sufficient data for groundwater taxa become available.
Our syncarids were collected from a fractured rock aquifer which had low hardness, low-organic matter water, contrasting to some previous studies in which biota were collected from karstic aquifers with much greater water hardness. Hardness is a major factor affecting the toxicity of metals, particularly zinc and chromium, to organisms [41,42,43], and hardness correction has been used to compare organism sensitivity in ecological risk assessments. Interestingly, using the hardness correction of Warne et al. [44], the syncarids tested were the most Zn-sensitive of the stygobitic crustaceans tested to date. No hardness correction algorithm is available for Cr(VI). With likely differences in the hardness of water between aquifer types, it is critical that aquifer types are considered in risk assessments and decision making for subterranean ecosystems and when setting environmental quality standards.
The toxicity of all toxicants increased with exposure period, with decreases in EC values of up to two orders of magnitude between 96 h and 14 days. The significance of such short-term exposures in groundwater ecosystems, with the exception of perhaps karst systems (see [34]), is debatable because of the slow groundwater flow rates in most aquifers. Hose et al. [16] showed that the relative toxicities of As, Cr and Zn were consistent between 96 h, 14-day and 28-day exposures, but with up to a six-fold decrease in ECx values between 14 and 28 days. Di Lorenzo et al. [10] recommend the reporting 96-h LCx values for the purposes of comparison between studies and the existing ecotoxicological databases, but recommend that tests be extended well beyond 96 h. Unfortunately, due to the longevity and low reproductive rates of stygobitic species, continuing tests for durations that are relevant to life cycles of the organisms tested may be difficult. For this reason, incipient LC50 values (sensu [45,46]) would be useful and should be a direction of future research.
The accumulation of both As and Zn appeared greatest in the region at the anterior of the thoracic region. Previous studies using decapods have shown high concentrations of both As [47] and Zn [48,49] in the hepatopancreas, which is consistent with the location of the thoracic hotspot in the syncarids, although we recognise that it is difficult to determine the specific organs with this method, and the uncertainty of the organ location on these small animals. Cresswell et al. [49] suggest that Zn is transported from the gill to the hepatopancreas for detoxication and/or metabolism. Cresswell et al. [49] also report high concentrations of Zn in the antennal gland, which is an important excretion site in crustaceans and which are well developed in the Bathynellacea [50]. However, this was not the case for Zn in this study, although this may explain the high concentration of As in that region. It is unclear which tissues in the posterior of the abdomen are causing the high Zn, but the distribution of As throughout the length of the abdomen may occur in variety of tissues of systems that extend through the syncarid abdomen [51]. Alternatively, it may be a result of water uptake through the abdomen as done by some crustaceans (sensu [52]) but it is unclear whether syncarids do this. Both White and Rainbow [48] and Cresswell et al. [49] report high Zn concentrations in the eye of decapods (associated with zinc metalloenzymes used in vision [53]), which was clearly not the case for our blind (eyeless) stygobitic syncarid.
The body concentration of As in the syncarids was far lower than that recorded for Zn, which reflects the relative abundance of these elements in the environment and most biological systems [54]. The concentration of As in the unexposed syncarids was 2.8 µg As/g and similar to unexposed amphipods and Daphnia (<5 µg As/g [55]). The concentrations of As in unexposed N. rhenorhodanensis [32] were below detection limit; however, the mean concentration of As in amphipods exposed to 0.1 mg As/L for 10 days (22.6 µg As/g) [32] was of a similar magnitude to the concentrations in syncarids (19 µg As/g) exposed to 0.15 mg As/L for a similar period in this study. The arsenic accumulation data suggest that the syncarids have no ability to regulate As uptake, with the body concentration for syncarids exposed to 0.15 mg As/L being significantly greater than those in the syncarids from controls, as well as mortality occurring at that concentration after 14 days. Arsenic accumulation by the syncarids did not increase significantly as exposure concentration increased beyond 0.22 mg As/L. It is unlikely that this reflects an ability to regulate As uptake at these concentrations since there was no evidence at the lower concentrations, and this has not been reported elsewhere. Rather, the lack of significant increase in body concentrations beyond 0.22 mg As/L is likely a statistical artefact caused by large variability in internal concentrations within the 0.22 mg As/L treatment, as evidenced by the size of the error bars in Figure 1A.
The background concentration of Zn in the syncarids was around 228 µg Zn/g which was similar to that for N. montellianus (113 µg Zn/g [38]). The accumulation of Zn by syncarids exposed to 1 mg Zn/L was significantly greater than syncarids in controls and exposed to lower concentrations, as seen in Figure 1B. This trend differs from that observed for N. montellianus, for which significant accumulation of Zn did not occur until they were exposed to 10 mg Zn /L [38], but this difference may reflect the greater hardness of the water used in that study (150–165 mg CaCO3/L), and the expected reduced bioavailability and uptake of metals by organisms with increasing hardness [56,57].
Our results suggest an ability of syncarids to regulate Zn at concentrations below 1 mg Zn/L. When considered alongside the mortality data, the increase in accumulation from the 0.38 to the 11 mg Zn/L exposures coincides approximately with a threshold change in mortality after 14 days. Rainbow and Luoma [58] detail several models of metal (particularly Zn) uptake in crustaceans, and that observed here for the syncarids most closely fits that common in the Decapoda (e.g., [59,60]), which might be expected given the phylogenetic proximity of the decapods and syncarids [61]. In this model, Zn concentration in the animals is regulated by excreting Zn until a threshold exposure concentration is reached beyond which the animal is unable to continue regulating their internal concentration [58]. Relatively little metal is stored in detoxified forms [59] and the contribution of Zn adsorbed passively to the exoskeleton to the total body concentration is small [59]. Above the threshold concentration, Zn is accumulated proportionally to the exposure concentration [58] and binds at sites where it will interfere with normal metabolic functioning, leading to toxicity [59]. While for some aquatic organisms the uptake of some metals from solution plateaus at high concentrations, this is has not been reported for invertebrates (e.g., [62]). The lack of apparent increase in mean body concentrations at exposure concentrations above 1 mg Zn/L may be a consequence of the large variability in body concentrations in those animals, which obscures an increasing trend.
Understanding the ecological significance of laboratory response data to population level effects in crustaceans in the field is difficult [63]. Our syncarids are no different, with the ecological significance of their bioaccumulation of Zn and As unclear. The accumulation of Zn by gammarid amphipods at concentrations as low as 0.3 mg Zn/L effected reproduction and energy utilisation in females [64]. At that concentration, the number of aborted broods increased, and the reduced energy absorption and reallocation of that energy away from reproduction resulted in smaller sized offspring [64]. The reproductive strategies of bathynellid syncarids are poorly known, but like other stygobiotic crustaceans they are expected to have low metabolic rates, reproductive rates and small brood sizes (i.e., k-strategy traits), a likely consequence of the low energy groundwater environment (Humphreys 2006). What is the effect of reducing energy absorption and fecundity in a population of organisms that are already at the lower extremes of such rates? Kammenga et al. [65] showed no difference in sensitivity of k-strategist and r-strategist nematodes to cadmium, but whether this holds true for other taxa is unclear.
Maltby and Naylor [64] predict that the smaller offspring resulting from zinc exposure will take longer to reach maturity and may reproduce at a smaller size, which typically means reduced fecundity, which is exacerbated by the increase in aborted broods. The net effect is expected to thus be reduced recruitment and population decline. Importantly for syncarids that feed on detrital matter, bacteria and fungi [66], changes in microbial assemblages, particularly fungal biomass, may also occur at relatively low Zn and As concentrations. Lategan et al. [67] observed 50% declines in biomass of groundwater fungi at 4.4 mg As/L and 0.61 mg Zn/L, thereby exacerbating potential sublethal effects on feeding and energy assimilation. In groundwaters where contamination is typically long-term and difficult to remediate, these impacts will be persistent, and likely lead to a decrease in population viability. The broader consequences of declining invertebrate populations in aquifers is the loss of ecosystem services [6], including changes to aquifer hydraulic properties [68].
Chromium, zinc and arsenic were acutely toxic to the stygobitic bathynellid syncarid. The accumulation of Zn was regulated at exposure concentrations below 1 mg Zn/L, above which body concentrations increased, leading to increased mortality. Arsenic was not regulated and was accumulated by the syncarids at all concentrations above the control. These are the first published toxicity data for syncarids and show them among the most sensitive of stygobitic crustaceans so far tested, likely in part due to the low hardness of the groundwater from the aquifer they inhabit and in which they were tested. The ecological significance of the toxicant accumulation and mortality may be significant given the consequent population effects and low capacity for stygobitic populations to recover.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4441/11/12/2508/s1, Figure S1: Dose response curves for bathynellid syncarids exposed to Arsenic (A, B), Chromium (C, D) and Zinc (E, F), for 96 h (A, C, E) and 14 days (B, D, F), Table S1: Fitted dose response curve parameters.
Author Contributions
Conceptualization, G.C.H. and M.J.L.; methodology, G.C.H., M.J.L., and R.S.; formal analysis, K.S., R.S. and G.C.H.; investigation, all; resources, G.C.H. and R.S.; writing—original draft preparation, G.C.H.; writing—review and editing, all; supervision, G.C.H. and M.J.L.; project administration, G.C.H.; funding acquisition, G.C.H. and R.S.
Funding
Bioaccumulation analyses were conducted at the Australian Nuclear Science and Technology Organisation through the support of grants ALNGRA10099 and AINGRA09097P. This project was also supported by CRC CARE project 1-1-08-06/7 and NSW Environmental Trust project 2005/RD/0108.
Acknowledgments
The authors are grateful for the assistance of Ashleigh Keast with the arsenic tests and Sarah Stephenson with field collections. The insightful comments of two anonymous reviewers improved this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Alley, W.M.; Healy, R.W.; LaBaugh, J.W.; Reilly, T.E. Flow and storage in groundwater systems. Science 2002, 296, 1985. [Google Scholar] [CrossRef] [PubMed]
- Morris, B.L.; Lawrence, A.R.L.; Chilton, P.J.C.; Adams, B.; Calow, R.C.; Klinck, B.A. Groundwater and Its Susceptibility to Degradation: A Global Assessment of the Problem and Options for Management; United Nations Environment Programme: Nairobi, Kenya, 2003; p. 126. [Google Scholar]
- Humphreys, W.F. Aquifers: The ultimate groundwater-dependent ecosystems. Aust. J. Bot. 2006, 54, 115–132. [Google Scholar] [CrossRef]
- Camacho, A.I.; Valdecasas, A.G. Global diversity of syncarids (Syncarida; Crustacea) in freshwater. In Freshwater Animal Diversity Assessment; Balian, E.V., Lévêque, C., Segers, H., Martens, K., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 257–266. [Google Scholar]
- Hose, G.C. Response to Humphreys’ (2007) Comments on Hose GC (2005) Assessing the Need for Groundwater Quality Guidelines for Pesticides Using the Species Sensitivity Distribution Approach. Hum. Ecol. Risk Assess. 2007, 13, 241–246. [Google Scholar] [CrossRef]
- Griebler, C.; Avramov, M.; Hose, G. Groundwater ecosystems and their services—Current status and potential risks. In Atlas of Ecosystem Services—Drivers, Risks, and Societal Responses; Schröter, M., Bonn, A., Klotz, S., Seppelt, R., Baessler, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 197–203. [Google Scholar]
- Hose, G.C.; Asmyhr, M.G.; Cooper, S.J.B.; Humphreys, W.F. Down under Down Under: Austral groundwater life. In Austral Ark: The State of Wildlife in Australia and New Zealand; Stow, A., Maclean, N., Holwell, G.I., Eds.; Cambridge University Press: Cambridge, UK, 2015; pp. 512–536. [Google Scholar]
- Castaño-Sánchez, A.; Hose, G.; Reboleira, A. Ecotoxicological effects of anthropogenic stressors in subterranean organisms: A review. Chemosphere 2019, in press. [Google Scholar] [CrossRef]
- Hose, G.C. Assessing the Need for Groundwater Quality Guidelines for Pesticides Using the Species Sensitivity Distribution Approach. Hum. Ecol. Risk Assess. 2005, 11, 951–966. [Google Scholar] [CrossRef]
- Di Lorenzo, T.; Di Marzio, W.D.; Fiasca, B.; Galassi, D.M.P.; Korbel, K.; Iepure, S.; Pereira, J.L.; Reboleira, A.S.P.S.; Schmidt, S.I.; Hose, G.C. Recommendations for ecotoxicity testing with stygobiotic species in the framework of groundwater environmental risk assessment. Sci. Total Environ. 2019, 681, 292–304. [Google Scholar] [CrossRef]
- Di Lorenzo, T.; Di Marzio, W.D.; Spigoli, D.; Baratti, M.; Messana, G.; Cannicci, S.; Galassi, D.M.P. Metabolic rates of a hypogean and an epigean species of copepod in an alluvial aquifer. Freshw. Biol. 2015, 60, 426–435. [Google Scholar] [CrossRef]
- Little, J.; Schmidt, D.J.; Cook, B.D.; Page, T.J.; Hughes, J.M. Diversity and phylogeny of south-east Queensland Bathynellacea. J. Aust. J. Zool. 2016, 64, 36–47. [Google Scholar] [CrossRef]
- Perina, G.; Camacho, A.I.; Huey, J.; Horwitz, P.; Koenders, A. New Bathynellidae (Crustacea) taxa and their relationships in the Fortescue catchment aquifers of the Pilbara region, Western Australia. Syst. Biodivers. 2019, 17, 148–164. [Google Scholar] [CrossRef]
- Camacho, A.I.; Mas-Peinado, P.; Dorda, B.A.; Casado, A.; Brancelj, A.; Knight, L.R.F.D.; Hutchins, B.; Bou, C.; Perina, G.; Rey, I. Molecular tools unveil an underestimated diversity in a stygofauna family: A preliminary world phylogeny and an updated morphology of Bathynellidae (Crustacea: Bathynellacea). Zool. J. Linn. Soc. 2017, 183, 70–96. [Google Scholar] [CrossRef]
- Korbel, K.L.; Hose, G.C. Habitat, water quality, seasonality, or site? Identifying environmental correlates of the distribution of groundwater biota. Freshw. Sci. 2015, 34, 329–343. [Google Scholar] [CrossRef]
- Hose, G.C.; Symington, K.; Lott, M.J.; Lategan, M.J. The toxicity of arsenic(III), chromium(VI) and zinc to groundwater copepods. Environ. Sci. Pollut. Res. 2016, 23, 18704–18713. [Google Scholar] [CrossRef] [PubMed]
- Fruchter, J. Peer Reviewed: In-Situ Treatment of Chromium-Contaminated Groundwater. Environ. Sci. Technol. 2002, 36, 464A–472A. [Google Scholar] [CrossRef] [PubMed]
- Scheeren, P.J.H.; Koch, R.O.; Buisman, C.J.N.; Barnes, L.J.; Versteegh, J.H. New biological treatment plant for heavy metal contaminated groundwater. In EMC ’91: Non-Ferrous Metallurgy—Present and Future; Springer: Dordrecht, The Netherlands, 1991; pp. 403–416. [Google Scholar]
- Smedley, P.L.; Kinniburgh, D.G. A review of the source, behaviour and distribution of arsenic in natural waters. Appl. Geochem. 2002, 17, 517–568. [Google Scholar] [CrossRef]
- Edmunds, W.M.; Shand, P. Groundwater Baseline Quality. In Natural Groundwater Quality; Edmunds, W.M., Shand, P., Eds.; Blackwell Publishing: Oxford, UK, 2009; pp. 1–21. [Google Scholar]
- Klockenkämper, R.; von Bohlen, A. Elemental Analysis of Environmental Samples by Total Reflection X-Ray Fluorescence: A Review. X-ray Spectrom. 1996, 25, 156–162. [Google Scholar] [CrossRef]
- Farías, S.S.; Casa, V.A.; Vázquez, C.; Ferpozzi, L.; Pucci, G.N.; Cohen, I.M. Natural contamination with arsenic and other trace elements in ground waters of Argentine Pampean Plain. Sci. Total Environ. 2003, 309, 187–199. [Google Scholar] [CrossRef]
- Siegele, R.; Cohen, D.D.; Dytlewski, N. The ANSTO high energy heavy ion microprobe. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. Atoms 1999, 158, 31–38. [Google Scholar] [CrossRef]
- Ryan, C.G. Developments in Dynamic Analysis for quantitative PIXE true elemental imaging. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. Atoms 2001, 181, 170–179. [Google Scholar] [CrossRef]
- Ryan, C.G.; Jamieson, D.N.; Churms, C.L.; Pilcher, J.V. A new method for on-line true-elemental imaging using PIXE and the proton microprobe. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. Atoms 1995, 104, 157–165. [Google Scholar] [CrossRef]
- Ritz, C.; Streibig, J.C. Bioassay analysis using R. J. Stat. Softw. 2005, 12, 1–22. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Hyne, R.V.; Pablo, F.; Julli, M.; Markich, S.J. Influence of water chemistry on the acute toxicity of copper and zinc to the cladoceran Ceriodaphnia cf dubia. Environ. Toxicol. Chem. 2005, 24, 1667–1675. [Google Scholar] [CrossRef] [PubMed]
- Hahn, H.J. The GW-Fauna-Index: A first approach to a quantitative ecological assessment of groundwater habitats. Limnol. Ecol. Manag. Inland Waters 2006, 36, 119–137. [Google Scholar] [CrossRef]
- Notenboom, J.; Cruys, K.; Hoekstra, J.; Vanbeelen, P. Effect of Ambient Oxygen Concentration Upon the Acute Toxicity of Chlorophenols and Heavy-Metals to the Groundwater Copepod Parastenocaris-Germanica (Crustacea). Ecotoxicol. Environ. Saf. 1992, 24, 131–143. [Google Scholar] [CrossRef]
- Lima, A.; Curtis, C.; Hammermeister, D.; Markee, T.; Northcott, C.E.; Brooke, L.T. Acute and chronic toxicities of arsenic(III) to fathead minnows, flagfish, daphnids, and an amphipod. Arch. Environ. Contam. Toxicol. 1984, 13, 595–601. [Google Scholar] [CrossRef]
- Canivet, V.; Chambon, P.; Gibert, J. Toxicity and bioaccumulation of arsenic and chromium in epigean and hypogean freshwater macroinvertebrates. Arch. Environ. Contam. Toxicol. 2001, 40, 345–354. [Google Scholar] [CrossRef]
- Di Marzio, W.D.; Castaldo, D.; Pantani, C.; Di Cioccio, A.; Di Lorenzo, T.; Saenz, M.E.; Galassi, D.M.P. Relative Sensitivity of Hyporheic Copepods to Chemicals. Bull. Environ. Contam. Toxicol. 2009, 82, 488–491. [Google Scholar] [CrossRef]
- Reboleira, A.S.; Abrantes, N.; Oromí, P.; Gonçalves, F. Acute Toxicity of Copper Sulfate and Potassium Dichromate on Stygobiont Proasellus: General Aspects of Groundwater Ecotoxicology and Future Perspectives. Water Air Soil Pollut. 2013, 224, 1–9. [Google Scholar] [CrossRef]
- Fargasova, A. Toxicity of Metals on Daphnia magna and Tubifex tubifex. Ecotoxicol. Environ. Saf. 1994, 27, 210–213. [Google Scholar] [CrossRef]
- Ewell, W.S.; Gorsuch, J.W.; Kringle, R.O.; Robillard, K.A.; Spiegel, R.C. Simultaneous evaluation of the acute effects of chemicals on seven aquatic species. Environ. Toxicol. Chem. 1986, 5, 831–840. [Google Scholar] [CrossRef]
- Meinel, W.; Krause, R. Zur Korrelation zwischen Zink und verschiedenen pH–Werten in in her toxischen Wirkung auf einige Grundwasser–Organismen. Z. Für Angew. Zool. 1988, 75, 159–182. [Google Scholar]
- Coppellotti Krupa, O.; Toniello, V.; Guidolin, L. Niphargus and Gammarus from karst waters: First data on heavy metal (Cd, Cu, Zn) exposure in a biospeleology laboratory. Subterr. Biol. 2004, 2, 33–41. [Google Scholar]
- Lazorchak, J.M.; Smith, M.E.; Haring, H.J. Development and validation of a Daphnia magna four-day survival and growth test method. Environ. Toxicol. Chem. 2009, 28, 1028–1034. [Google Scholar] [CrossRef] [PubMed]
- Daam, M.A.; Silva, E.; Leitao, S.; Trindade, M.J.; Cerejeira, M.J. Does the actual standard of 0.1 mu g/L overestimate or underestimate the risk of plant protection products to groundwater ecosystems? Ecotoxicol. Environ. Saf. 2010, 73, 750–756. [Google Scholar] [CrossRef] [PubMed]
- Paulauskis, J.D.; Winner, R.W. Effects of water hardness and humic acid on zinc toxicity to Daphnia magna Straus. Aquat. Toxicol. 1988, 12, 273–290. [Google Scholar] [CrossRef]
- Pawlisz, A.V.; Kent, R.A.; Schneider, U.A.; Jefferson, C. Canadian water quality guidelines for chromium. Environ. Toxicol. Water Qual. 1997, 12, 123–183. [Google Scholar] [CrossRef]
- Park, E.J.; Jo, H.J.; Jung, J. Combined effects of pH, hardness and dissolved organic carbon on acute metal toxicity to Daphnia magna. J. Ind. Eng. Chem. 2009, 15, 82–85. [Google Scholar] [CrossRef]
- Warne, M.; Batley, G.; van Dam, R.; Chapman, J.; Fox, D.; Hickey, C.; Stauber, J. Revised Method for Deriving Australian and New Zealand Water Quality Guideline Values for Toxicants—Update of 2015 Version; Australian and New Zealand Governments and Australian State and Territory Governments: Canberra, Australia, 2018; p. 48.
- Sprague, J.B. Measurement of pollutant toxicity to fish I. Bioassay methods for acute toxicity. Water Res. 1969, 3, 793–821. [Google Scholar] [CrossRef]
- Avramov, M.; Schmidt, S.I.; Griebler, C. A new bioassay for the ecotoxicological testing of VOCs on groundwater invertebrates and the effects of toluene on Niphargus inopinatus. Aquat. Toxicol. 2013, 130, 1–8. [Google Scholar] [CrossRef]
- Williams, G.; West, J.M.; Snow, E.T. Total arsenic accumulation in yabbies (Cherax destructor clark) exposed to elevated arsenic levels in Victorian gold mining areas, Australia. Environ. Toxicol. Chem. 2008, 27, 1332–1342. [Google Scholar] [CrossRef]
- White, S.L.; Rainbow, P.S. Regulation of zinc concentration by Palaemon elegans (Crustacea: Decapoda): Zinc flux and effects of temperature, zinc concentration and moulting. Mar. Ecol. Prog. Ser. 1984, 16, 135–147. [Google Scholar] [CrossRef]
- Cresswell, T.; Simpson, S.L.; Mazumder, D.; Callaghan, P.D.; Nguyen, A.P. Bioaccumulation Kinetics and Organ Distribution of Cadmium and Zinc in the Freshwater Decapod Crustacean Macrobrachium australiense. Environ. Sci. Technol. 2015, 49, 1182–1189. [Google Scholar] [CrossRef] [PubMed]
- Steenken, B.; Schminke, H.K. Ultrastructure of maxillary gland of Antrobathynella stammeri (Syncarida, Malacostraca). J. Morphol. 1996, 228, 107–117. [Google Scholar] [CrossRef]
- Smith, G. On the Anaspidacea, Living and Fossil. J. Cell Sci. 1909, s2-53, 489–578. [Google Scholar]
- Fox, H.M. Anal and Oral Intake of Water by Crustacea. J. Exp. Biol. 1952, 29, 583. [Google Scholar] [CrossRef]
- White, S.L.; Rainbow, P.S. Zinc flux in Palaemon elegans (Crustacea: Decapoda): Moulting, individual variation and tissue distribution. Mar. Ecol. Prog. Ser. 1984, 19, 153–166. [Google Scholar] [CrossRef]
- Smith, K.S.; Huyck, H. An overview of the abundance, relative mobility, bioavailabilty, and human toxicity of metals. In The Environmental Geochemistry of Mineral Deposits: Part, A; Processes, Techniques, and Health Issues; Plumlee, G., Logsdon, J., Eds.; Society for Economic Geologists: Littleton, CO, USA, 1999; Volume 6A, pp. 29–70. [Google Scholar]
- Spehar, R.L.; Fiandt, J.T.; Anderson, R.L.; DeFoe, D. Comparative toxicity of arsenic compounds and their accumulation in invertebrates and fish. Arch. Environ. Contam. Toxicol. 1980, 9, 53–63. [Google Scholar] [CrossRef]
- Winner, R.W.; Gauss, J.D. Relationship between chronic toxicity and bioaccumulation of copper, cadmium and zinc as affected by water hardness and humic acid. Aquat. Toxicol. 1986, 8, 149–161. [Google Scholar] [CrossRef]
- Markich, S.; Brown, P.; Batley, G.; Apte, S.; Stauber, J. Incorporating metal speciation and bioavailability into water quality guidelines for protecting aquatic ecosystems. Aust. J. Ecotoxicol. 2001, 7, 109–122. [Google Scholar]
- Rainbow, P.S.; Luoma, S.N. Metal toxicity, uptake and bioaccumulation in aquatic invertebrates—Modelling zinc in crustaceans. Aquat. Toxicol. 2011, 105, 455–465. [Google Scholar] [CrossRef]
- Rainbow, P.S. Trace metal bioaccumulation: Models, metabolic availability and toxicity. Environ. Int. 2007, 33, 576–582. [Google Scholar] [CrossRef]
- Vijayram, K.; Geraldine, P. Toxicology. Regulation of Essential Heavy Metals (Cu, Cr, and Zn) by the Freshwater Prawn Macrobrachium malcolmsonii (Milne Edwards). Bull. Environ. Contam. Toxicol. 1996, 56, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Koenemann, S.; Jenner, R.A.; Hoenemann, M.; Stemme, T.; von Reumont, B.M. Arthropod phylogeny revisited, with a focus on crustacean relationships. Arthropod Struct. Dev. 2010, 39, 88–110. [Google Scholar] [CrossRef] [PubMed]
- Luoma, S.N.; Rainbow, P.S. Metal. Contamination in Aquatic Environments: Science and Lateral Management; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar] [CrossRef]
- Kuhn, A.; Munns, W.R., Jr.; Serbst, J.; Edwards, P.; Cantwell, M.G.; Gleason, T.; Pelletier, M.C.; Berry, W. Evaluating the ecological significance of laboratory response data to predict population-level effects for the estruarine amphipod Ampelisca abdita. Environ. Toxicol. Chem. 2002, 21, 865–874. [Google Scholar] [CrossRef] [PubMed]
- Maltby, L.; Naylor, C. Preliminary Observations on the Ecological Relevance of the Gammarus ‘Scope for Growth’ Assay: Effect of Zinc on Reproduction. Funct. Ecol. 1990, 4, 393–397. [Google Scholar] [CrossRef]
- Kammenga, J.E.; Van Gestel, C.A.M.; Bakker, J. Toxicology. Patterns of sensitivity to cadmium and pentachlorophenol among nematode species from different taxonomic and ecological groups. Arch. Environ. Contam. Toxicol. 1994, 27, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Wellborn, G.A.; Witt, J.D.S.; Cothran, R.D. Chapter 31—Class Malacostraca, Superorders Peracarida and Syncarida. In Thorp and Covich’s Freshwater Invertebrates, 4th ed.; Thorp, J.H., Rogers, D.C., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 781–796. [Google Scholar] [CrossRef]
- Lategan, M.J.; Klare, W.; Kidd, S.; Hose, G.C.; Nevalainen, H. The unicellular fungal tool RhoTox for risk assessments in groundwater systems. Ecotoxicol. Environ. Saf. 2016, 132, 18–25. [Google Scholar] [CrossRef]
- Hose, G.C.; Stumpp, C. Architects of the underworld: Bioturbation by groundwater invertebrates influences aquifer hydraulic properties. Aquat. Sci. 2019, 81, 20. [Google Scholar] [CrossRef]
Figure 1.
Mean concentration (±SE) between exposure concentration and mean body concentration in bathynellid syncarids exposed to (A) As and (B) Zn. Common lower case letters indicate no significant difference (p > 0.05) between those treatments.
Figure 1.
Mean concentration (±SE) between exposure concentration and mean body concentration in bathynellid syncarids exposed to (A) As and (B) Zn. Common lower case letters indicate no significant difference (p > 0.05) between those treatments.

Figure 2.
Images of As accumulation in bathynellid syncarids exposed to As in aqueous solution. Mean concentrations of specimens shown were (A) 12 ppm, (B) 2 ppm, and (C) 28 ppm.
Figure 2.
Images of As accumulation in bathynellid syncarids exposed to As in aqueous solution. Mean concentrations of specimens shown were (A) 12 ppm, (B) 2 ppm, and (C) 28 ppm.

Figure 3.
Images of Zn accumulation in bathynellid syncarids exposed to Zn in aqueous solution. Mean concentrations of specimens shown were (A) 250 ppm, (B) 6400 ppm, and (C) 8000 ppm.
Figure 3.
Images of Zn accumulation in bathynellid syncarids exposed to Zn in aqueous solution. Mean concentrations of specimens shown were (A) 250 ppm, (B) 6400 ppm, and (C) 8000 ppm.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Physicochemical variables and concentrations of common metals in the groundwater at the Somersby (NSW, Australia) collection sites (from [16]).
Table 1.
Physicochemical variables and concentrations of common metals in the groundwater at the Somersby (NSW, Australia) collection sites (from [16]).
| Water Quality Variable | Units | Concentration |
|---|---|---|
| pH | 4.2–5.6 | |
| Conductivity | µS/cm | 131–195 |
| Dissolved Oxygen | % Saturation | 59–83 |
| Hardness | mg/L as CaCO3 | 25–44 |
| Total Organic Carbon | mg/L | 3–13 |
| As | mg/L | <0.01 |
| Cr | mg/L | 0.03 |
| Zn | mg/L | 0.02 |
Table 2.
Toxicity of As(III), Cr(VI) and Zn to bathynellid syncarids. EC10 and EC50 values indicate the concentration (mg/L) effecting 10% and 50% of the test population, respectively. Values in parentheses indicate 95% fiducial limits, followed by the total number of animals tested (N).
Table 2.
Toxicity of As(III), Cr(VI) and Zn to bathynellid syncarids. EC10 and EC50 values indicate the concentration (mg/L) effecting 10% and 50% of the test population, respectively. Values in parentheses indicate 95% fiducial limits, followed by the total number of animals tested (N).
| As(III) | Cr(VI) | Zn(II) | |||||
|---|---|---|---|---|---|---|---|
| Test Duration | ECx | Estimate | 95% Confidence Interval | Estimate | 95% Confidence Interval | Estimate | 95% Confidence Interval |
| N = 74 | N = 71 | N = 190 | |||||
| 96 h | 10 | 0.17 | (−0.04–0.38) | 0.11 | (−0.07–0.28) | 0.78 | (0.30–1.26) |
| 50 | 1.99 | (0.79–3.19) | 1.83 | (−0.10–3.77) | 3.61 | (2.56–4.66) | |
| 14 days | 10 | 0.01 | (−0.02–0.04) | 0.004 | (−0.010–0.018) | 0.21 | (0.02–0.39) |
| 50 | 0.25 | (0.07–0.44) | 0.51 | (−0.07–1.09) | 1.77 | (1.11–2.43) | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hose, G.C.; Symington, K.; Lategan, M.J.; Siegele, R. The Toxicity and Uptake of As, Cr and Zn in a Stygobitic Syncarid (Syncarida: Bathynellidae). Water 2019, 11, 2508. https://doi.org/10.3390/w11122508
AMA Style
Hose GC, Symington K, Lategan MJ, Siegele R. The Toxicity and Uptake of As, Cr and Zn in a Stygobitic Syncarid (Syncarida: Bathynellidae). Water. 2019; 11(12):2508. https://doi.org/10.3390/w11122508
Chicago/Turabian StyleHose, Grant C., Katelyn Symington, Maria J. Lategan, and Rainer Siegele. 2019. "The Toxicity and Uptake of As, Cr and Zn in a Stygobitic Syncarid (Syncarida: Bathynellidae)" Water 11, no. 12: 2508. https://doi.org/10.3390/w11122508
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.