Diatom Biodiversity in Karst Springs of Mediterranean Geographic Areas with Contrasting Characteristics: Islands vs Mainland

 , , , , , , and
, , , , , , and 
Abstract
:1. Introduction
2. Materials and Methods
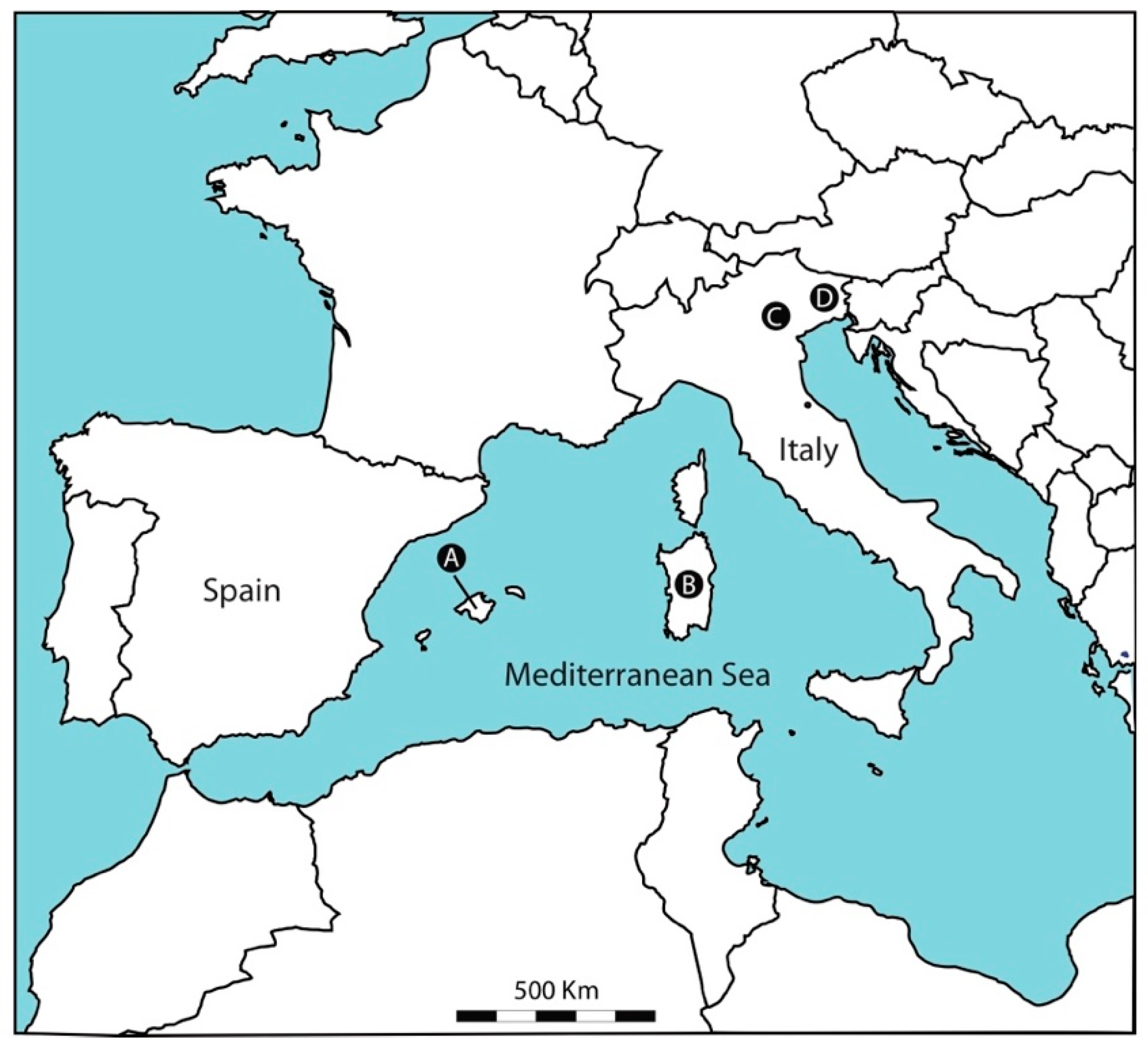
2.1. Study Sites
2.2. Sampling
2.3. Laboratory Analyses
2.4. Data Processing and Statistical Analyses
3. Results
3.1. Environmental Variables
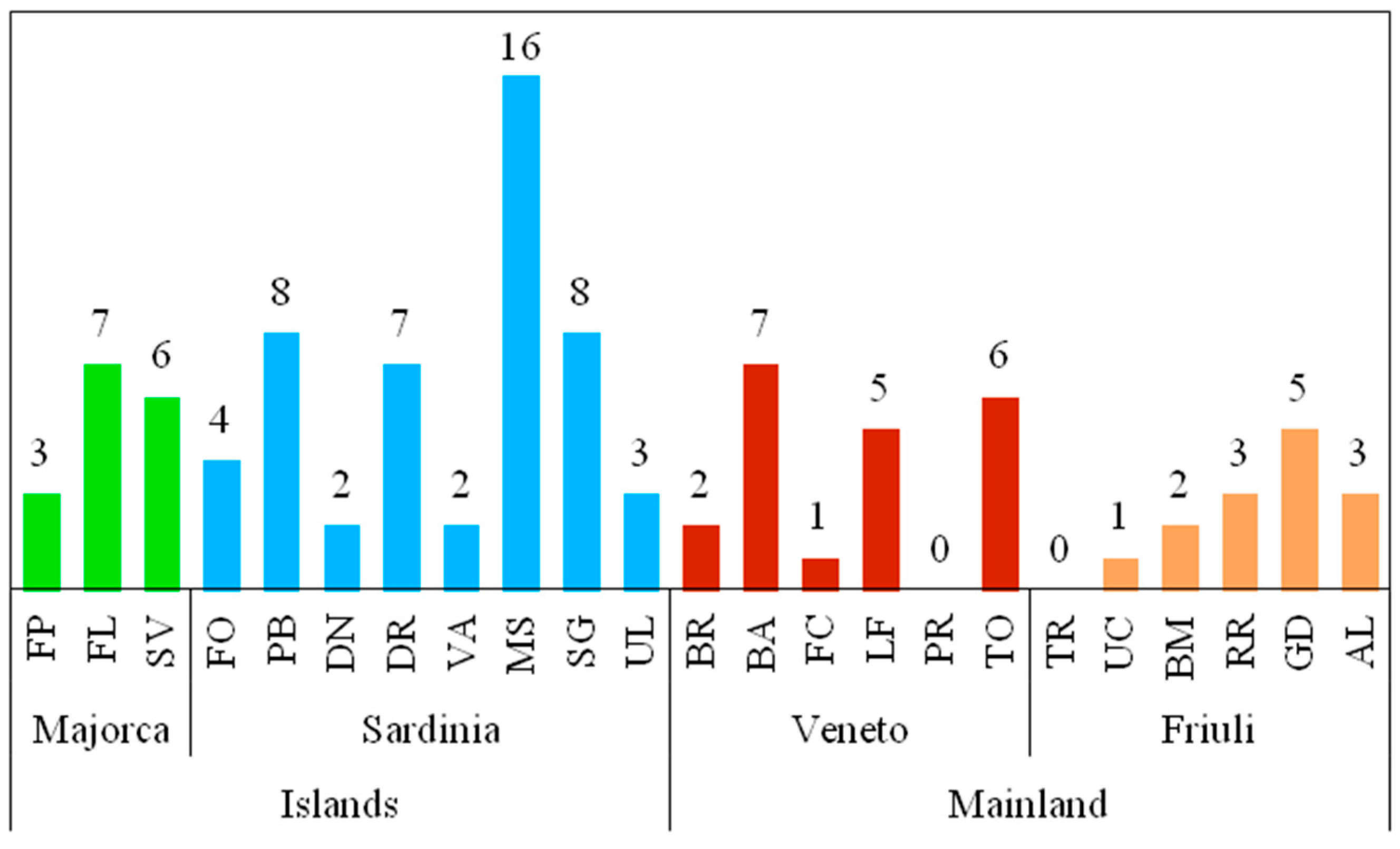
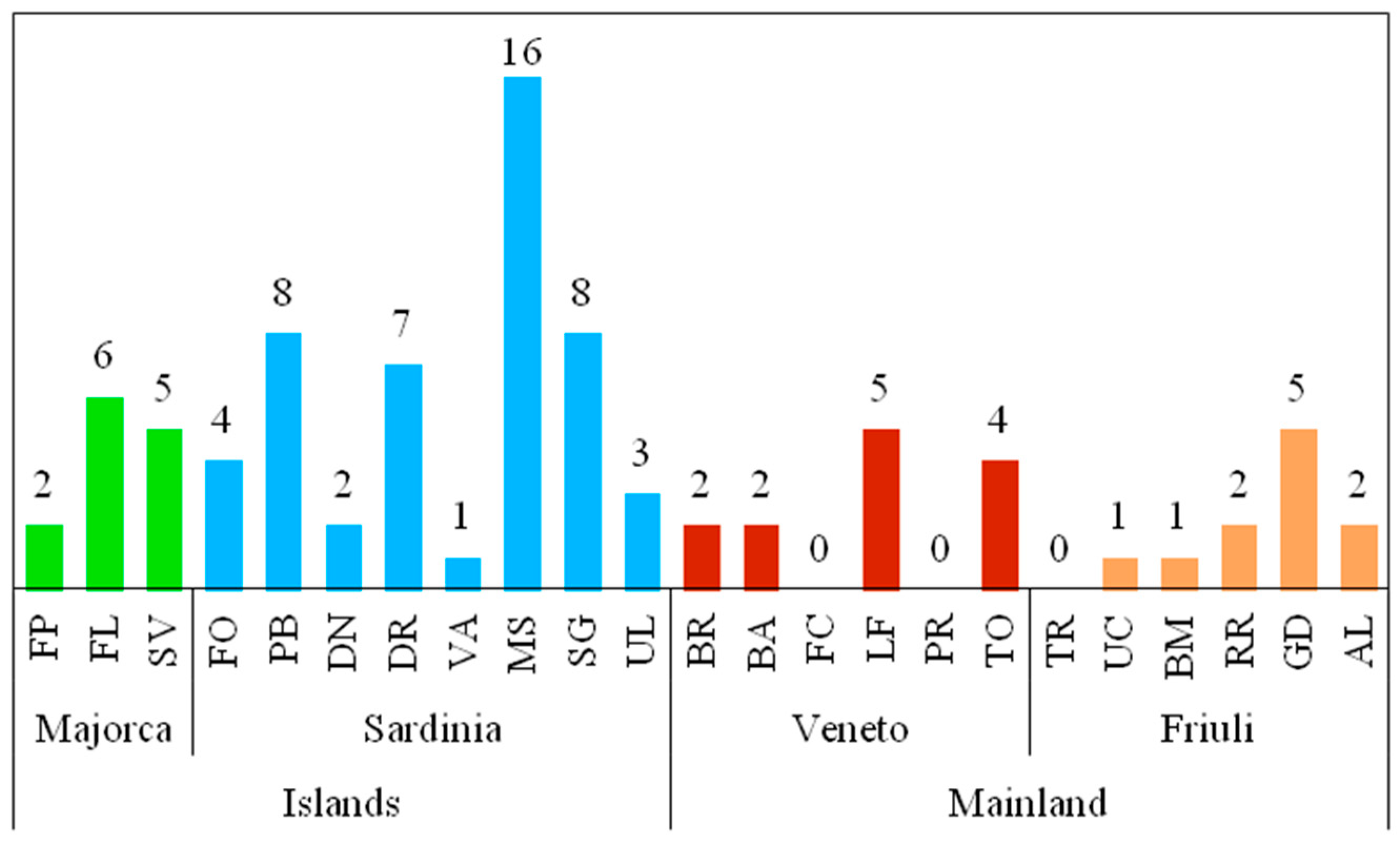
3.2. Species Composition and Structure of Diatom Assemblages
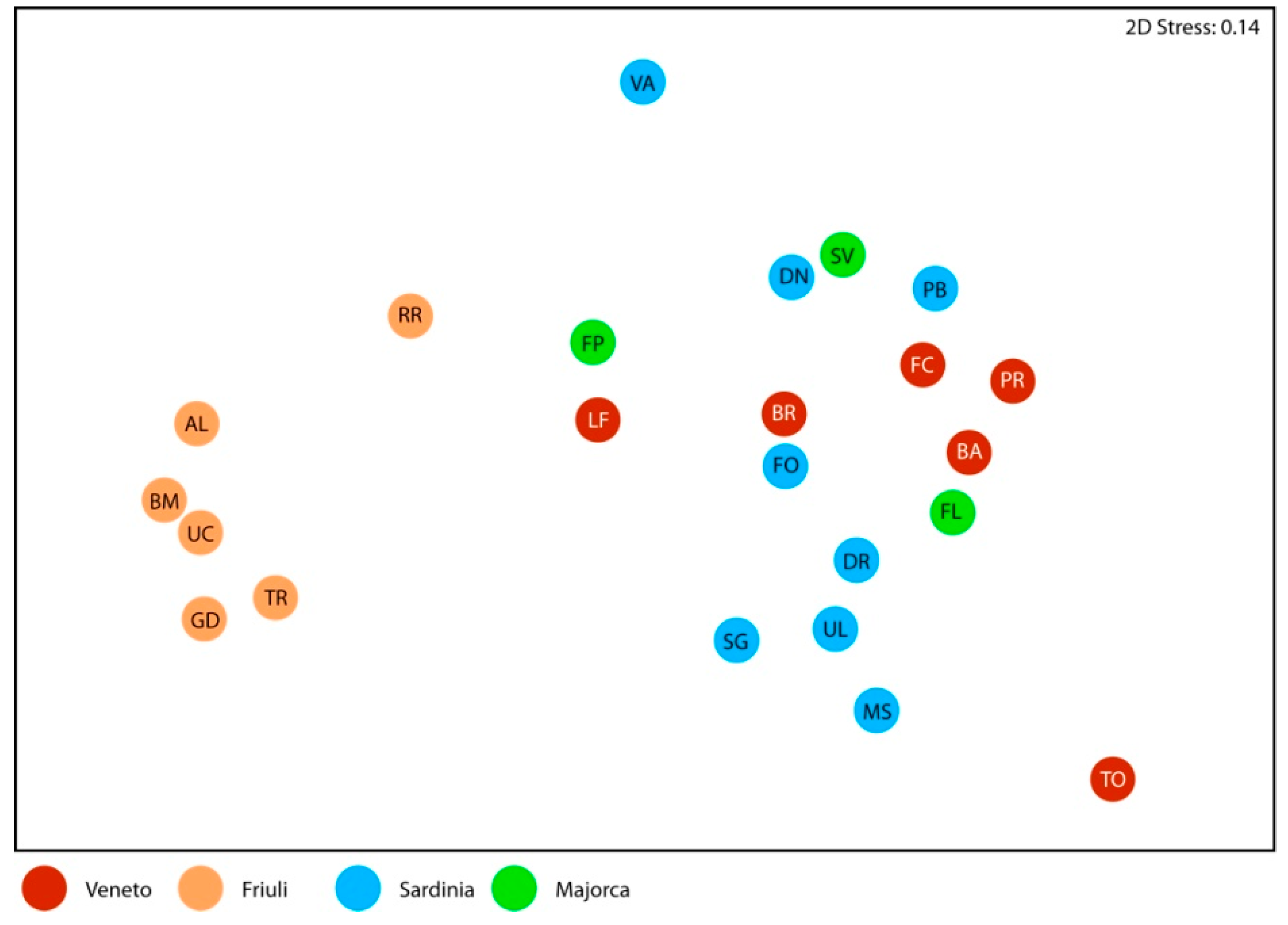
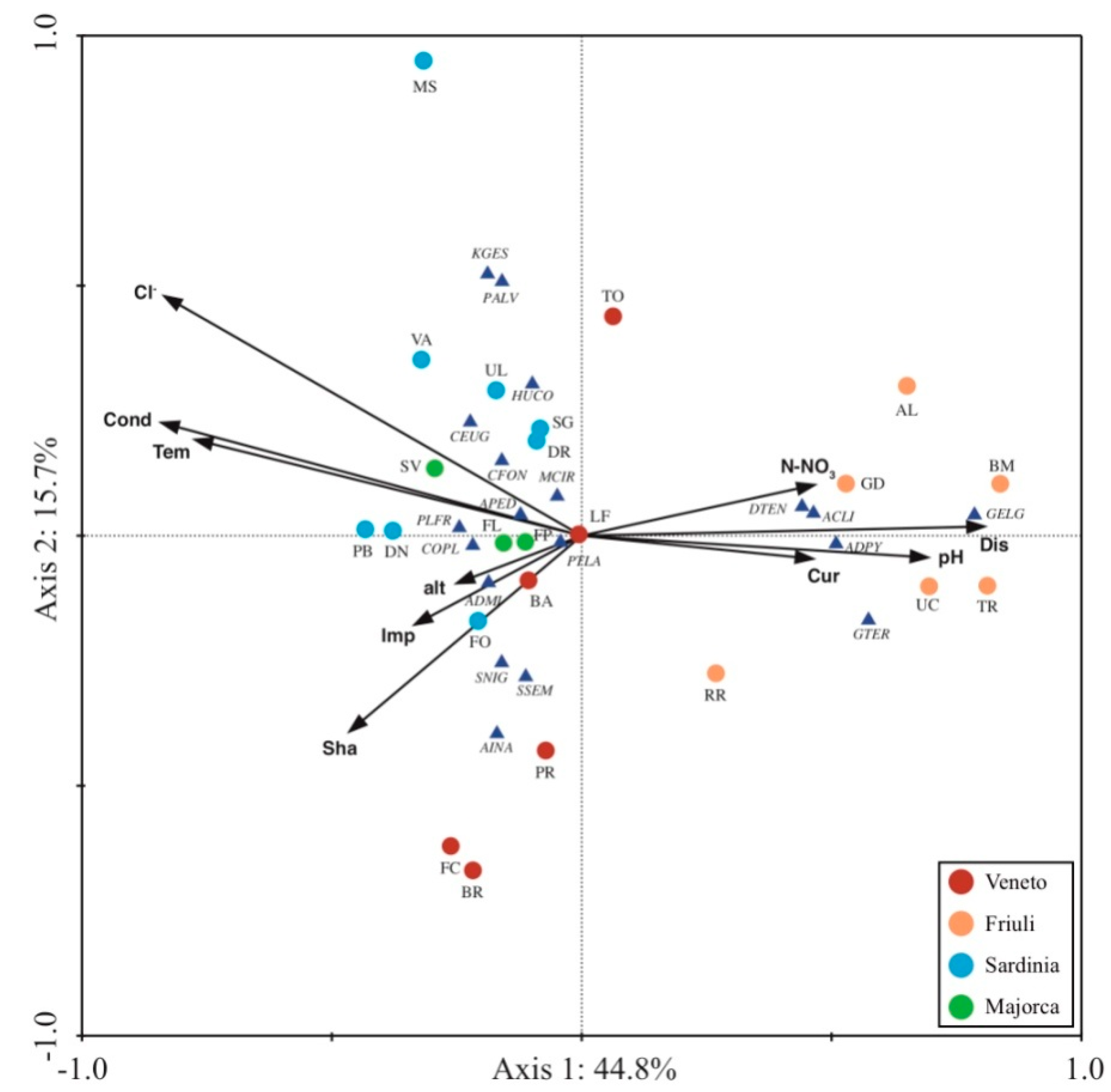
3.3. Comparison among Diatom Assemblages
3.4. Relationships with Environmental Variables
4. Discussion
4.1. Main Characteristics of the Diatom Assemblages
4.2. Structure of the Diatom Assemblages
4.3. Comparison of Diatom Assemblages from Islands and Mainland
4.4. Relationships Diatoms-Environmental Variables
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Parise, M.; Gabrovsek, F.; Kaufmann, G.; Ravbar, N. Recent advances in karst research: From theory to fieldwork and applications. In Advances in Karst Research: Theory, Fieldwork and Application; Parise, M., Gabrovsek, F., Kaufmann, G., Ravbar, N., Eds.; Geological Society: London, UK, 2018; Special Publications 466; pp. 1–24. [Google Scholar]
- Fiorillo, F.; Stevanović, Z. Introductory editorial thematic issue: Mediterranean karst hydrogeology. Environ. Earth Sci. 2015, 74, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Bakalowicz, M.; Fleury, P.; Dörfliger, N.; Seidel, J.L. Coastal karst aquifers in Mediterranean regions. A valuable ground water resource in complex aquifers. In Proceedings of the Technologia de la Intrusion de Agua de Mar en Acuiferos Costeros: Paises Mediterraneos (TIAC ’03), Alicante, Spain, 11–14 March 2003; López Geta, J.A., de Dios Gómez, J., de la Orden, V., Ramos, G., Rodríguez, L., Eds.; Instituto Geológico y Minero de España: Madrid, Spain, 2003; pp. 125–128. [Google Scholar]
- Bonacci, O. Karst landscape ecohydrology. In Proceedings of the International Symposium on Water Management and Hydraulic Engineering, Ohrid, Macedonia, 1–5 September 2009; pp. 781–790. [Google Scholar]
- Zelnik, I.; Balanč, T.; Toman, M.J. Diversity and Structure of the tychoplankton diatom community in the limnocrene spring Zelenci (Slovenia) in relation to environmental factors. Water 2018, 10, 361. [Google Scholar] [CrossRef] [Green Version]
- Maiolini, B.; Carolli, M.; Silveri, L. Ephemeroptera, Plecoptera and Trichoptera in springs in Trentino (south-eastern Alps). J. Limnol. 2011, 70 (Suppl. 1), 122–133. [Google Scholar] [CrossRef]
- Taxböck, L.; Linder, H.P.; Cantonati, M. To what extent are Swiss springs refugial habitats for sensitive and endangered diatom taxa? Water 2017, 9, 967. [Google Scholar] [CrossRef] [Green Version]
- Cantonati, M.; Füreder, L.; Gerecke, R.; Jüttner, I.; Cox, E.J. Crenic habitats, hotspots for freshwater biodiversity conservation: Toward an understanding of their ecology. Freshw. Sci. 2012, 312, 463–480. [Google Scholar] [CrossRef]
- Meyer, A.; Meyer, E.I.; Meyer, C. Lotic communities of two small temporary karstic stream systems (East Westphalia, Germany) along a longitudinal gradient of hydrological intermittency. Limnologica 2003, 33, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Daly, D.; Dassargues, A.; Drew, D.; Dunne, S.; Goldscheider, N.; Neale, S.; Popescu, I.C.; Zwahlen, F. Main concepts of the “European approach” to karst-groundwater-vulnerability assessment and mapping. Hydrogeol. J. 2002, 10, 340–345. [Google Scholar] [CrossRef] [Green Version]
- Bolle, H.J. Mediterranean Climate: Variability and Trends, 1st ed.; Springer: Berlin, Germany, 2003; pp. 1–372. [Google Scholar]
- Intergovernmental Panel on Climate Change—IPCC. Climate Change 2007: The Physical Science Basis; Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate, Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2007; pp. 1–996. [Google Scholar]
- Spaɫek, K.; Procków, J. New, rare and constant habitats for endangered aquatic plant communities: The importance of microhabitats for global biodiversity. Diversity 2019, 11, 96. [Google Scholar] [CrossRef] [Green Version]
- Herman, J.W.; Culver, D.C.; Salzman, J. Groundwater ecosystems and the service of water purification. Stan. Environ. Law J. 2001, 20, 479–495. [Google Scholar]
- Goldscheider, N. A holistic approach to groundwater protection and ecosystem services in karst terrains. AQUA Mundi 2012, Am06046, 117–124. [Google Scholar] [CrossRef]
- Poulíčková, A.; Hájková, P.; Křenková, P.; Hájek, M. Distribution of diatoms and bryophytes on linear transects through spring fens. Nova Hedwigia 2004, 78, 411–424. [Google Scholar] [CrossRef]
- Cantonati, M.; Gerecke, R.; Bertuzzi, E. Springs of the Alps–sensitive ecosystems to environmental change: From biodiversity assessments to long-term studies. Hydrobiologia 2006, 562, 59–96. [Google Scholar] [CrossRef]
- Mogna, M.; Cantonati, M.; Andreucci, F.; Angeli, N.; Berta, G.; Miserere, L. Diatom communities and vegetation of springs in the south-western Alps. Acta Bot. Croat. 2015, 742, 265–285. [Google Scholar] [CrossRef] [Green Version]
- Werum, M. Die Kieselalgengesellschaften in Quellen. Abhängigkeit von Geologie und anthropogener Beeinflussung in Hessen (Bundesrepublik Deutschland); Hessisches Landesamt fur Umwelt und Geologie: Wiesbaden, Germany, 2001; p. 273.
- Rott, E.; Pfister, P.; Pipp, E.; Pall, K.; Binder, N.; Ortler, K. Indikationslisten für Aufwuchsalgen in Fließgewässern. Teil 2; Bundesministerium für Land-u. Forstwirtschaft, Wasserwirtschaftskataster: Wien, Austria, 1999; p. 248. [Google Scholar]
- Menegalija, T.; Kosi, G. Distribution of diatoms in springs in the Julian Alps (NW Slovenia). Nat. Slov. 2008, 101, 21–37. [Google Scholar]
- Žutinić, P.; Petrić, I.; Gottstein, S.; Udovič, M.G.; Borojević, K.K.; Kamberović, J.; Kolda, A.; Plenković-Moraj, A.; Ternjej, I. Microbial mats as shelter microhabitat for amphipods in an intermittent karstic spring. Knowl. Manag. Aquat. Ecosyst. 2018, 419, 7. [Google Scholar] [CrossRef]
- Dedić, A.; Plenković-Moraj, A.; Borojević, K.K.; Hafner, D. The first report on periphytic diatoms on artificial and natural substrate in the karstic spring Bunica, Bosnia and Herzegovina. Acta Bot. Croat. 2015, 742, 393–406. [Google Scholar] [CrossRef] [Green Version]
- Angeli, N.; Cantonati, M.; Spitale, D.; Lange-Bertalot, H. A comparison between diatom assemblages in two groups of carbonate, low-altitude springs with different levels of anthropogenic disturbances. Fottea 2010, 101, 115–128. [Google Scholar] [CrossRef] [Green Version]
- Cantonati, M.; Bertuzzi, E.; Scalfi, A. CRENODAT (biodiversity assessment and integrity evaluation of springs of Trentino (Italian Alps) and long-term ecological research): Project design and preliminary results. In Integrated Watershed Management: Perspectives and Problems; Beheim, E., Rajwar, G.S., Haigh, M., Krecek., J., Eds.; Springer, Capital Publishing Company: New Delhi, India, 2010; pp. 121–132. [Google Scholar]
- Delgado, C.; Ector, L.; Novais, M.H.; Blanco, S.; Hoffmann, L.; Pardo, I. Epilithic diatoms of springs and spring–fed streams in Majorca Island (Spain) with the description of a new diatom species Cymbopleura margalefii sp. nov. Fottea 2013, 132, 87–104. [Google Scholar] [CrossRef] [Green Version]
- Dell’Uomo, A. Chorological analysis of the algal flora of “Su Gologone” spring in Sardinia. In Proceedings of the International Symposium on Biogeographical Aspects of Insularity, Rome, Italy, 18–22 May 1987; Accademia Nazionale dei Lincei: Rome, Italy, 1990; pp. 417–427. [Google Scholar]
- Lai, G.G.; Padedda, B.M.; Wetzel, C.E.; Lugliè, A.; Sechi, N.; Ector, L. Valutazione della qualità ambientale della sorgente carsica Su Gologone (Sardegna centro-orientale) mediante l’impiego delle diatomee epilitiche: Primi risultati. Biol. Ambient. 2015, 291, 37–45. [Google Scholar]
- Lai, G.G.; Padedda, B.M.; Wetzel, C.E.; Lugliè, A.; Sechi, N.; Ector, L. Epilithic diatom assemblages and environmental quality of the Su Gologone karst spring (central-eastern Sardinia, Italy). Acta Bot. Croat. 2016, 751, 129–143. [Google Scholar] [CrossRef] [Green Version]
- Lai, G.G.; Ector, L.; Lugliè, A.; Sechi, N.; Wetzel, C.E. Sellaphora gologonica sp. nov. (Bacillariophyta, Sellaphoraceae), a new diatom species from a Mediterranean karst spring (Sardinia, Italy). Phytotaxa 2018, 3562, 145–157. [Google Scholar] [CrossRef]
- Lai, G.G.; Ector, L.; Wetzel, C.E.; Sechi, N.; Lugliè, A.; Padedda, B.M. Periphytic diatoms of the Mediterranean karst spring Sa Vena (Su Gologone system, Sardinia, Italy): Relationships with environmental variables and effects of an extreme flash flood. Inland Waters 2018, 83, 284–293. [Google Scholar] [CrossRef]
- Lai, G.G.; Padedda, B.M.; Ector, L.; Wetzel, C.E.; Lugliè, A.; Cantonati, M. Mediterranean karst springs: Diatom biodiversity hotspots under the pressure of hydrological fluctuation and nutrient enrichment. Plant Biosyst. 2019. [Google Scholar] [CrossRef]
- Llobera, M.; Ferriol, A. Seasonal fluctuations in the chemistry of limestone springs from the Tramuntana Mountain Range (Mallorca, Spain). Verh. Int. Ver. Limnol. 1994, 25, 1445–1448. [Google Scholar] [CrossRef]
- Available online: http://sgi.isprambiente.it/GeositiWeb/default.aspx (accessed on 10 October 2019).
- Poldini, L.; Oriolo, G.; Vidali, M.; Tomasella, M.; Stoch, F.; Orel, G. Manuale Degli Habitat del Friuli Venezia Giulia. Strumento a Supporto Della Valutazione D’impatto Ambientale (VIA), Ambientale Strategica (VAS) e di Incidenza Ecologica (VIEc). Regione Autonoma Friuli Venezia Giulia—Direzione Centrale Ambiente e Lavori Pubblici—Serv. VIA, Univ. Degli STUDI di Trieste—Dip. Biologia. Available online: http://www.regione.fvg.it/ambiente/manuale/home.htm (accessed on 30 July 2019).
- Cantonati, M.; Spitale, D. The role of environmental variables in structuring epiphytic and epilithic diatom assemblages in springs and streams of the Dolomiti Bellunesi National Park (south-eastern Alps). Fundam. Appl. Limnol. Arch. Hydrobiol. 2009, 1742, 117–133. [Google Scholar] [CrossRef]
- CEN. Water Quality—Guidance Standard for the Routine Sampling and Pretreatment of Benthic Diatoms from Rivers; European Standard EN 13946; European Committee for Standardization: Brussels, Belgium, 2003; pp. 1–14. [Google Scholar]
- Agenzia per la Protezione Dell’ambiente e per i Servizi Tecnici/Istituto di Ricerca Sulle Acque—Consiglio Nazionale delle Ricerche. Metodi Analitici per le Acque; Rapporti 29/2003; I.G.E.R. srl. APAT: Rome, Italy, 2003; ISBN 88-448-0083-7. Available online: http://www.isprambiente.gov.it/it/pubblicazioni/manuali-e-linee-guida/metodi-analitici-per-le-acque (accessed on 7 October 2017).
- APHA. Standard Methods for the Examination of Water and Wastewater, 20th ed.; APHA, AWWA & WEF, American Public Health Association: Washington, DC, USA, 2000; pp. 1–1220. [Google Scholar]
- Strickland, J.D.H.; Parsons, T.R. A Practical Handbook of Seawater Analysis, Bulletin 167, 2nd ed.; Fisheries Research Board of Canada: Ottawa, ON, Canada, 1972; pp. 1–328. [Google Scholar]
- Istituto Superiore per la Protezione e la Ricerca Ambientale. Metodi Biologici per le Acque Superficali Interne; Manuali e Linee Guida 111/2014; Istituto Superiore per la Protezione e la Ricerca Ambientale: Rome, Italy, 2003. Available online: http://admin.isprambiente.gov.it/it/pubblicazioni/manuali-e-linee-guida/metodi-biologici-per-le-acque-superficiali-interne (accessed on 10 June 2017).
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae. Teil 1–4; Gustav Fischer Verlag: Stuttgart, Germany; New York, NY, USA, 1991. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae. English and French Translation of the Keys Teil 5; Spektrum Akademischer Verlag: Berlin/Heidelberg, Germany, 2000; pp. 1–311. [Google Scholar]
- Lange-Bertalot, H.; Cavacini, P.; Tagliaventi, N.; Alfinito, S. Diatoms of Sardinia. Rare and 76 New Species in Rock Pools and Other Ephemeral Waters; Iconographia Diatomologica 12; A.R.G. Gantner Verlag, KG: Ruggell, Germany, 2003; pp. 1–438. [Google Scholar]
- Lange-Bertalot, H.; Hofmann, G.; Werum, M.; Cantonati, M. Freshwater Benthic Diatoms of Central Europe: Over 800 Common Species Used in Ecological Assessment; English edition with updated taxonomy and added species; Koeltz Botanical Books: Schmitten, Oberreifenberg, Germany, 2017; pp. 1–942. [Google Scholar]
- Wojtal, A.Z. Species Composition and Distribution of Diatom Assemblages in Spring Waters from Various Geological Formations in Southern Poland; Bibliotheca Diatomologica 59; J. Cramer: Stuttgart, Germany, 2013; pp. 1–436. [Google Scholar]
- Żelazna-Wieczorek, J. Diatom Flora in Springs of Lódź Hills (Central Poland). Biodiversity, Taxonomy, and Temporal Changes of Episammic Diatom Assemblages in Springs Affected by Human Impact, Diatom Monographs 13; A.R.G. Gantner: Ruggell, Liechtenstein, Germany, 2011; pp. 1–419. [Google Scholar]
- Cantonati, M.; Lange-Bertalot, H. Geissleria gereckei sp. nov. (Bacillariophyta) from leaf-litter covered stones of very shaded carbonate mountain springs with extremely low discharge. Phycol. Res. 2009, 57, 171–177. [Google Scholar] [CrossRef]
- Novais, M.H.; Blanco, S.; Hlúbiková, D.; Falasco, E.; Gomà, J.; Delgado, C.; Ivanov, P.; Àcs, E.; Morais, M.; Hoffmann, L.; et al. Taxonomic revision and biogeography of the Gomphonema rosenstockianum and G. tergestinum species complex. Fottea 2009, 9, 257–274. [Google Scholar] [CrossRef] [Green Version]
- Cejudo-Figueiras, C.; Morales, E.A.; Wetzel, C.E.; Blanco, S.; Hoffmann, L.; Ector, L. Analysis of the type of Fragilaria construens var. subsalina (Bacillariophyceae) and description of two morphologically related taxa from Europe and the United States. Phycologia 2011, 501, 67–77. [Google Scholar] [CrossRef]
- Wetzel, C.E.; Ector, L.; Van de Vijver, B.; Compère, P.; Mann, D.G. Morphology, typification and critical analysis of some ecologically important small naviculoid species (Bacillariophyta). Fottea 2015, 152, 203–234. [Google Scholar] [CrossRef] [Green Version]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication, 1st ed.; University of Illinois Press: Urbana, IL, USA, 1949; pp. 1–117. [Google Scholar]
- Pielou, E.C. Ecological Diversity; Wiley: New York, NY, USA, 1975; pp. 1–165. [Google Scholar]
- Lecointe, C.; Coste, M.; Prygiel, J. “Omnidia”: Software for taxonomy, calculation of diatom indices and inventories management. Hydrobiologia 1993, 269, 509–513. [Google Scholar] [CrossRef]
- Student. The probable error of a mean. Biometrika 1908, 61, 1–25. [Google Scholar]
- Fisher, R.A. Applications of “Student’s’ distribution”. Metron 1925, 5, 90–104. [Google Scholar]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: http://www.r-project.org/index.html (accessed on 20 September 2019).
- Van Dam, H.; Mertens, A.; Sinkeldam, J. A coded checklist and ecological indicator values of freshwater diatoms from The Netherlands. Netherland J. Aquat. Ecol. 1994, 28, 117–133. [Google Scholar]
- Kruskal, J.B. Nonmetric multidimensional scaling: A numerical method. Psychometrika 1964, 29, 115–129. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 1st ed.; Plymouth Marine Laboratory: Plymouth, UK, 1994; pp. 1–144. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial (Plymouth Routines in Multivariate Ecological Research); Plymouth Marine Laboratory: Plymouth, UK, 2006; pp. 1–190. [Google Scholar]
- Ter Braak, C.J.F. Canonical correspondence analysis: A new eigenvector technique for multivariate direct gradient analysis. Ecology 1986, 67, 1167–1179. [Google Scholar] [CrossRef] [Green Version]
- Hill, M.O.; Gauch, H.G., Jr. Detrended correspondence analysis: An improved ordination technique. Vegetatio 1980, 42, 47–58. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Šmilauer, P. CANOCO Reference Manual and Cano Draw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002; pp. 1–500. [Google Scholar]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 673, 345–366. [Google Scholar] [CrossRef]
- Bertrand, J.; Renon, J.-P.; Monnier, O. Les diatomées des sources du rebord karstique de la Beauce de la région orléanaise. Symbioses 1999, 1, 3–14. [Google Scholar]
- Wojtal, A.Z.; Sobczyk, Ł. The influence of substrates and physicochemical factors on the composition of diatom assemblages in karst springs and their applicability in water-quality assessment. Hydrobiologia 2012, 6951, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Niedermayr, R.; Schagerl, M. Structuring factors of the phytobenthos community along a mountain headwater (Kalkalpen National Park, Austria). Fundam. Appl. Limnol. 2010, 1772, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Cantonati, M.; Lange-Bertalot, H. Diatom biodiversity of springs in the Berchtesgaden National Park (northern Alps, Germany), with the ecological and morphological characterization of two species new to science. Diatom Res. 2010, 252, 251–280. [Google Scholar] [CrossRef]
- Cantonati, M.; Angeli, N.; Spitale, D.; Lange–Bertalot, H. A new Navicula (Bacillariophyta) species from low–elevation carbonate springs affected by anthropogenic disturbance. Fottea 2016, 162, 255–265. [Google Scholar] [CrossRef]
- Cantonati, M.; Lange-Bertalot, H.; Scalfi, A.; Angeli, N. Cymbella tridentina sp. nov. (Bacillariophyta), a crenophilous diatom from carbonate springs of the Alps. J. N. Am. Benthol. Soc. 2010, 293, 775–788. [Google Scholar] [CrossRef]
- Lai, G.G.; Padedda, B.M.; Wetzel, C.E.; Cantonati, M.; Sechi, N.; Lugliè, A.; Ector, L. Diatom assemblages from different substrates of the Casteldoria thermo-mineral spring (Northern Sardinia, Italy). Bot. Lett. 2019, 1661, 14–31. [Google Scholar] [CrossRef]
- Lai, G.G.; Beauger, A.; Wetzel, C.E.; Padedda, B.M.; Voldoire, O.; Lugliè, A.; Allain, E.; Ector, L. Diversity, ecology and distribution of benthic diatoms in thermo-mineral springs in Auvergne (France) and Sardinia (Italy). PeerJ 2019, 7, e7238. [Google Scholar] [CrossRef]
- Cantonati, M.; Rott, E.; Spitale, D.; Angeli, N.; Komárek, J. Are benthic algae related to spring types? Freshw. Sci. 2012, 31, 481–498. [Google Scholar] [CrossRef]
- Gerecke, R.; Cantonati, M.; Spitale, D.; Stur, E.; Wiedenbrug, S. The challenges of long-term ecological research in springs in the northern and southern Alps: Indicator groups, habitat diversity, and medium-term change. J. Limnol. 2011, 70 (Suppl. 1), 168–187. [Google Scholar] [CrossRef]
- Noga, T.; Rybak, M. First record of Pinnularia subinterrupta Krammer & Schroeter in Poland—A rare species in Europe. Biodivers. Res. Conserv. 2017, 451, 17–21. [Google Scholar]
- Moyá, G.; Ferriol, A.; Llobera, M. Chlorophyll contents in the freshwater vegetation of Serra de Tramuntana springs (Mallorca, Spain). Verh. Int. Ver. Limnol. 1991, 24, 2002–2006. [Google Scholar]
- Falasco, E.; Piano, E.; Bona, F. Guida al riconoscimento e all’ecologia delle principali diatomee fluviali dell’Italia nord occidentale. Biol. Ambient. 2013, 27, 1–292. [Google Scholar]
- Monnier, O.; Rimet, F.; Bey, M.; Chavaux, R.; Ector, L. Sur l’identité de Cocconeis euglypta Ehrenberg 1854 et C. lineata Ehrenberg 1843. Un approche par le sources historiques. Diatomania 2007, 11, 30–45. [Google Scholar]
- Blanco, S.; Cejudo-Figueiras, C.; Álvarez-Blanco, I.; Becares, E.; Hoffmann, L.; Ector, L. Atlas de las Diatomeas de la Cuenca del Duero, 1st ed.; Área de Publicaciones, Universidad de León: Leon, Spain, 2010; pp. 1–382. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spring Name | Code | Altitude (m.a.s.l.) | Latitude (N) | Longitude (E) | Type | Regime | Spring-Tapping |
|---|---|---|---|---|---|---|---|
| Majorca | |||||||
| Font des Pí | FP | 471 | 39°46′14″ | 2°48′26″ | R | P | X |
| Font de s′ Olla | FL | 56 | 39°45′23″ | 2°42′42″ | R | P | X |
| Son Vich | SV | 356 | 39°39′12″ | 2°32′08″ | R | P | X |
| Sardinia | |||||||
| Fruncu ‘e Oche | FO | 55 | 40°33′84″ | 009°40′67″ | R | P | X |
| Pubusinu | PB | 212 | 39°24′34″ | 008°31′39″ | R | P | X |
| San Giovanni-DN | DN | 190 | 39°20′13″ | 008°37′37″ | R | P | X |
| San Giovanni-DR | DR | 168 | 40°19′14″ | 009°36′55″ | R | P | X |
| Sa Varva | VA | 780 | 40°13′39″ | 009°12′31″ | R | S | |
| Sa Vena Manna-S | MS | 276 | 40°50′25″ | 008°48′20″ | R | P | X |
| Su Gologone-SVM | SG | 116 | 40°17′34″ | 009°29′80″ | L-R | P | |
| S′Ulidone | UL | 606 | 40°34′30″ | 009°38′35″ | R | P | X |
| Veneto | |||||||
| Brusaferri | BR | 675 | 45°35′57″ | 11°12′59″ | R-L | P | X |
| Buso delle Anguane | BA | 370 | 45°33′04″ | 11°17′07″ | R-L | P | X |
| Fonte del Coppo | FC | 183 | 45°28′39″ | 11°06′51″ | L-R | P | X |
| La Ferrara | LF | 640 | 45°32′59″ | 11°11′08″ | L-R | P | X |
| Prealba | PR | 445 | 45°37′34″ | 10°49′47″ | L-R | P | X |
| Torricelle | TO | 220 | 45°28′09″ | 11°00′16″ | R-L | P | X |
| Friuli | |||||||
| Torre | TR | 528 | 46°18′27″ | 13°16′21″ | R | P | |
| Uccea | UC | 1.122 | 46°20′02″ | 13°19′30″ | R | P | |
| Barman | BM | 652 | 46°20′21″ | 13°17′18″ | R | P | |
| Rio Researtico | RR | 454 | 46°22′22″ | 13°13′01″ | R | P | X |
| Goriuda | GD | 701 | 46°23′44″ | 13°26′8″ | R | P | |
| Alba | AL | 1.249 | 46°28′12″ | 13°13′48″ | R | P |
| Majorca | Sardinia | Veneto | Friuli | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Variable/Spring | FL | FP | SV | FO | PB | DN | DR | VA | MS | SG | UL | FC | BA | PR | TO | LF | BR | TR | UC | BM | RR | GD | AL |
| T [°C] | 15.7 | 12.5 | 15.1 | 15.1 | 15.9 | 14.2 | 13.4 | 13.2 | 16.4 | 13.0 | 9.5 | 13 | 12.2 | 11.9 | 12 | 12.3 | 10.3 | 8.7 | 8.3 | 6.7 | 10.8 | 13.1 | 7.3 |
| pH [units] | 7.25 | 6.65 | 6.91 | 7.62 | 7.80 | 8.10 | 7.74 | 6.98 | 7.49 | 7.52 | 7.20 | 7.83 | 7.9 | 7.71 | 7.6 | 7.2 | 7.2 | 8.32 | 8.24 | 8.71 | 8.38 | 8.47 | 8.40 |
| Conductivity [μS cm−1] | 612 | 416 | 796 | 339 | 571 | 478 | 439 | 760 | 1140 | 326 | 357 | 424 | 333 | 438 | 436 | 498 | 578 | 154 | 170 | 136 | 148 | 131 | 229 |
| Impacts [score] | 4 | 1 | 3 | 8 | 7 | 8 | 9 | 3 | 9 | 3 | 3 | 10 | 5 | 10 | 3 | 10 | 5 | 7 | 5 | 2 | 3 | 3 | 2 |
| Shading [score] | 5 | 2 | 4 | 3 | 2 | 3 | 2 | 4 | 1 | 3 | 3 | 5 | 2 | 2 | 2 | 3 | 4 | 1 | 1 | 3 | 1 | 3 | 1 |
| Discharge [L s−1] | 10–50 | 0.5–1 | 5–10 | 0.5–1 | <0.5 | <0.5 | <0.5 | 0.5–1 | <0.5 | 1–5 | <0.5 | <0.5 | <0.5 | 0.5–1 | <0.5 | <0.5 | 0.02 | >100 | 10–50 | >100 | 5–10 | 0.5–1 | >50 |
| Current velocity [score] | 4 | 2 | 3 | 3 | 2 | 2 | 2 | 3 | 2 | 3 | 3 | 3 | 2 | 2 | 2 | 3 | 2 | 4 | 4 | 4 | 4 | 1 | 4 |
| P-PO43− [µg L−1] | 1 | 1 | 1 | 14 | 7 | 8 | 161 | 8 | 66 | 10 | 9 | - | - | - | - | - | - | 0 | 10 | 0 | 10 | 20 | 0 |
| N-NO3− [µg L−1] | 225 | 134 | 231 | 371 | 460 | 309 | 2093 | 1549 | 3634 | 806 | 49 | 2257 | 1354 | 2257 | 5643 | 5869 | 1580 | 3000 | 2300 | 2300 | 2300 | 6200 | 3000 |
| Cl− [mg L−1] | 24.1 | 15.7 | 45.0 | 28.4 | 56.7 | 60.3 | 39.0 | 42.5 | 195.0 | 21.3 | 46.0 | 7.8 | 15.0 | 9.9 | 11.7 | 15.0 | 6.0 | 1.0 | 0.5 | 0.4 | 0.6 | 4.1 | 0.5 |
| Ca2+ [mg L−1] | 97.5 | 75.3 | 85.8 | 30.0 | 42.0 | 32.0 | 48.0 | 56.0 | 54.0 | 32.0 | 18.0 | - | 52.4 | - | - | 106.2 | 111.5 | 24.4 | 35.8 | 23.7 | 21.3 | 25.4 | 22.0 |
| Mg2+ [mg L−1] | 22.5 | 13.0 | 35.1 | 18.2 | 27.9 | 21.9 | 18.2 | 51.0 | 48.6 | 19.4 | 18.2 | - | 13.6 | - | - | 52.7 | 15.7 | 7.4 | 1.0 | 4.6 | 8.9 | 2.2 | 12.8 |
| Majorca | Sardinia | Veneto | Friuli | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Taxa | FP | FL | SV | FO | PB | DN | DR | VA | MS | SG | UL | BR | BA | FC | LF | PR | TO | TR | UC | BM | RR | GD | AL |
| Achnanthidium exile (Kützing) Heiberg | 9.2 | ||||||||||||||||||||||
| Achnanthidium lineare W. Smith | 1.7 | 0.9 | 0.6 | 0.8 | 7.7 | 2.4 | 0.5 | 0.5 | 5.4 | 7.0 | |||||||||||||
| Achnanthidium minutissimum (Kützing) Czarnecki | 76.6 | 7.9 | 64.6 | 16.4 | 35.1 | 31.2 | 5.3 | 4.3 | 0.9 | 3.6 | 1.0 | 43.3 | 9.8 | 31.4 | 69.2 | 15.7 | 4.4 | ||||||
| Achnanthidium pyrenaicum (Hustedt) Kobayasi | 6.1 | 2.5 | 7.2 | 52.6 | 74.9 | 30.0 | 6.3 | 63.4 | 22.9 | ||||||||||||||
| Amphora inariensis Krammer | 0.6 | 7.8 | 12.3 | 40.2 | 1.6 | 1.6 | |||||||||||||||||
| Amphora indistincta Levkov | 1.0 | 0.5 | 3.0 | 0.7 | 11.1 | 2.9 | |||||||||||||||||
| Amphora pediculus (Kützing) Grunow | 2.1 | 27.0 | 1.2 | 6.9 | 1.5 | 17.6 | 0.7 | 15.3 | 21.7 | 17.9 | 10.7 | 11.9 | 7.9 | 3.9 | 0.2 | 1.0 | 0.5 | 0.5 | |||||
| Caloneis fontinalis (Grunow) Cleve-Euler | 0.7 | 0.7 | 3.9 | 6.0 | 5.6 | 1.0 | 0.8 | ||||||||||||||||
| Cocconeis euglypta Ehrenberg | 0.9 | 14.3 | 7.7 | 6.1 | 7.5 | 12.1 | 11.1 | 1.7 | 0.2 | ||||||||||||||
| Cocconeis pseudolineata (Geitler) Lange-Bertalot | 1.6 | 46.2 | 0.5 | 7.2 | 0.5 | 9.4 | 0.5 | 1.0 | |||||||||||||||
| Cymbella affinis Kützing | 1.4 | 1.5 | 9.8 | ||||||||||||||||||||
| Cymbella tridentina Lange-Bertalot, M. Cantonati & A. Scalfi | 32.3 | ||||||||||||||||||||||
| Derticula tenuis Kützing | 0.2 | 8.0 | 0.5 | 7.0 | 0.5 | 2.0 | 1.5 | ||||||||||||||||
| Diploneis separanda Lange-Bertalot | 0.2 | 6.3 | 0.5 | 2.1 | 0.4 | ||||||||||||||||||
| Ellerbeckia arenaria (Moore ex Ralfs) R.M. Crawford | 5.3 | 0.7 | 1.5 | ||||||||||||||||||||
| Encyonema minutum (Hilse) D.G. Mann | 0.3 | 1.5 | 1.5 | 7.3 | |||||||||||||||||||
| Encyonema ventricosum (Agardh) Grunow | 0.5 | 11.1 | 2.0 | 4.4 | |||||||||||||||||||
| Eunotia arcubus Nörpel & Lange-Berlalot | 15.3 | ||||||||||||||||||||||
| Fallacia insociabilis (Krasske) D.G. Mann | 0.2 | 20.0 | |||||||||||||||||||||
| Fallacia muraloides (Hustedt) D.G. Mann | 5.7 | ||||||||||||||||||||||
| Fallacia sublucidula (Hustedt) D.G. Mann | 0.2 | 5.3 | |||||||||||||||||||||
| Gomphonema aff. cymbelliclinum Reichardt & Lange-Bertalot | 30.2 | ||||||||||||||||||||||
| Gomphonema elegantissimum Reichardt & Lange-Bertalot | 26.8 | 5.8 | 47.5 | 3.0 | |||||||||||||||||||
| Gomphonema micropus Kützing | 0.5 | 6.2 | 1.2 | 1.5 | 0.2 | ||||||||||||||||||
| Gomphonema rosenstockianum Lange-Bertalot & Reichardt | 0.9 | 3.7 | 18.6 | ||||||||||||||||||||
| Gomphonema sp.2 | 12.0 | ||||||||||||||||||||||
| Gomphonema subclavatum Grunow | 0.5 | 0.2 | 1.5 | 17.4 | |||||||||||||||||||
| Gomphonema tergestinum (Grunow) Fricke | 7.2 | 1.9 | 6.3 | ||||||||||||||||||||
| Gomphosphenia lingulatiformis (Lange-Bertalot & Reichardt) Lange-Bertalot | 32.7 | 0.6 | |||||||||||||||||||||
| Hannaea arcus (Ehrenberg) R.M. Patrick | 10.2 | ||||||||||||||||||||||
| Humidophila contenta (Grunow) Lowe, Kociolek et al. | 3.7 | 0.2 | 0.2 | 5.9 | 3.6 | 2.0 | 1.6 | 57.3 | |||||||||||||||
| Kolbesia gessneri (Hustedt) Aboal | 9.6 | 12.3 | 1.9 | ||||||||||||||||||||
| Meridion circulare (Greville) C. Agardh | 0.2 | 5.7 | 1.7 | 0.2 | 61.2 | 16.2 | 0.6 | 41.5 | 3.0 | ||||||||||||||
| Nitzschia frustulum (Kützing) Grunow | 7.4 | ||||||||||||||||||||||
| Nitzschia inconspicua Grunow | 0.7 | 17.4 | 0.9 | 0.2 | 0.5 | 2.1 | |||||||||||||||||
| Nitzschia recta Hantzsch ex Rabenhorst | 0.2 | 5.2 | |||||||||||||||||||||
| Odontidium mesodon (Ehrenberg) Kützing | 0.2 | 0.5 | 0.5 | 18.0 | 0.5 | 0.5 | |||||||||||||||||
| Planothidium frequentissimum (Lange-Bertalot) Lange-Bertalot | 9.7 | 3.2 | 4.5 | 28.6 | 1.2 | 2.5 | 1.8 | 16.0 | 1.3 | 0.7 | 1.3 | 0.6 | 8.0 | 2.3 | |||||||||
| Planothidium lanceolatum (Brébisson ex Kützing) Lange-Bertalot | 12.9 | 0.7 | 0.9 | 6.0 | 13.6 | 4.4 | 0.4 | 1.5 | 0.0 | 1.9 | 0.5 | 0.5 | |||||||||||
| Platessa conspicua (A. Mayer) Lange-Bertalot | 11.9 | ||||||||||||||||||||||
| Psammothidium grischunum (Wuthrich) Bukhtiyarova & Round | 7.7 | ||||||||||||||||||||||
| Pseudostaurosira alvareziae Cejudo-Figueras, E.A. Morales & Ector | 12.3 | 5.7 | |||||||||||||||||||||
| Rhoicosphenia abbreviata (C. Agardh) Lange-Bertalot | 1.8 | 0.2 | 5.9 | 0.9 | |||||||||||||||||||
| Sellaphora atomoides (Grunow) C.E. Wetzel & Van de Vijver | 1.7 | 1.4 | 20.8 | ||||||||||||||||||||
| Sellaphora nigri (De Notaris) C.E. Wetzel & Ector | 0.2 | 13.2 | 10.2 | 0.7 | 5.5 | 11.6 | 0.9 | 9.0 | 0.0 | 26.3 | 1.0 | ||||||||||||
| Sellaphora seminulum (Grunow) D.G. Mann | 2.3 | 1.3 | 8.2 | 0.6 | 0.0 | 21.7 | |||||||||||||||||
| Spring | R | H’ | J’ |
|---|---|---|---|
| Majorca | |||
| FP | 14 | 1.40 | 0.37 |
| FL | 19 | 3.19 | 0.75 |
| SV | 16 | 1.94 | 0.49 |
| Sardinia | |||
| FO | 27 | 3.77 | 0.79 |
| PB | 22 | 2.93 | 0.66 |
| DN | 18 | 2.21 | 0.53 |
| DR | 43 | 4.37 | 0.81 |
| VA | 8 | 1.44 | 0.48 |
| MS | 54 | 4.48 | 0.78 |
| SG | 37 | 4.05 | 0.78 |
| UL | 25 | 3.22 | 0.69 |
| Veneto | |||
| BR | 16 | 2.51 | 0.76 |
| BA | 21 | 2.90 | 0.81 |
| FC | 10 | 2.00 | 0.77 |
| LF | 20 | 1.49 | 0.45 |
| PR | 13 | 2.47 | 0.78 |
| TO | 16 | 1.83 | 0.58 |
| Friuli | |||
| TR | 20 | 1.94 | 0.47 |
| UC | 26 | 1.43 | 0.31 |
| BM | 35 | 1.91 | 0.39 |
| RR | 19 | 2.80 | 0.67 |
| GD | 34 | 2.09 | 0.43 |
| AL | 22 | 2.76 | 0.62 |
| Groups | R Statistic | p Significance Level |
|---|---|---|
| Global effect | 0.610 | 0.001 |
| Pairwise tests: | ||
| Veneto vs Sardinia | 0.278 | 0.008 |
| Veneto vs Majorca | 0.068 | 0.333 |
| Veneto vs Friuli | 0.898 | 0.002 |
| Sardinia vs Majorca | 0.052 | 0.358 |
| Sardinia vs Friuli | 0.928 | 0.001 |
| Majorca vs Friuli | 0.963 | 0.012 |
| Groups of Sites | Species | Code | IndVal | p-Value |
|---|---|---|---|---|
| 1 (Friuli) | Achnanthidium pyrenaicum | ADPY | 0.975 | 0.001 |
| Achnanthidium lineare | ACLI | 0.950 | 0.002 | |
| Gomphonema elegantissimum | GELG | 0.816 | 0.005 | |
| Gomphonema tergestinum | GTER | 0.707 | 0.018 | |
| 2 (Majorca + Sardinia) | Cocconeis euglypta | CEUG | 0.850 | 0.003 |
| 3 (Veneto) | Amphora inariensis | AINA | 0.911 | 0.001 |
| Sellaphora seminulum | SSEM | 0.801 | 0.007 | |
| Sellaphora nigri | SNIG | 0.791 | 0.027 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, G.G.; Burato, S.; Padedda, B.M.; Zorza, R.; Pizzul, E.; Delgado, C.; Lugliè, A.; Cantonati, M. Diatom Biodiversity in Karst Springs of Mediterranean Geographic Areas with Contrasting Characteristics: Islands vs Mainland. Water 2019, 11, 2602. https://doi.org/10.3390/w11122602
Lai GG, Burato S, Padedda BM, Zorza R, Pizzul E, Delgado C, Lugliè A, Cantonati M. Diatom Biodiversity in Karst Springs of Mediterranean Geographic Areas with Contrasting Characteristics: Islands vs Mainland. Water. 2019; 11(12):2602. https://doi.org/10.3390/w11122602
Chicago/Turabian StyleLai, Giuseppina G., Sara Burato, Bachisio M. Padedda, Raffaella Zorza, Elisabetta Pizzul, Cristina Delgado, Antonella Lugliè, and Marco Cantonati. 2019. "Diatom Biodiversity in Karst Springs of Mediterranean Geographic Areas with Contrasting Characteristics: Islands vs Mainland" Water 11, no. 12: 2602. https://doi.org/10.3390/w11122602