1. Introduction
Hydrocarbon pollution is a widespread phenomenon that affects human health and the environment, including air, water, and soil [
1]. Hydrocarbons of petroleum origin, despite being essential energy resources and one of the raw materials needed for different types of industries [
2], are classified as priority pollutants [
3]. Many of them, such as Polycyclic Aromatic Hydrocarbons (PAHs), are recalcitrant and highly dangerous, as they can be hemotoxic, carcinogenic and teratogenic [
3,
4,
5,
6,
7]. Accordingly, over the decades, the awareness to protect the environment has increased, especially now that pollution is recognized as one of the most severe and urgent issues society has to face. The sources of environmental contamination caused by hydrocarbons are different, such as accidents in the transport of fuel by ships and tankers, leaks from underground tanks and service stations, oil extraction and processing operations, release of oily waste generated by industries that use oil in the production of plastics, solvents, pharmaceuticals and cosmetics [
8,
9]. The cleaning up of these contaminants from the environment is mandatory and can be reached by using physical-chemical or biological strategies [
1,
10]. Biological approaches have shown several advantages compared to traditional physical-chemical treatments, being more cost effective and allowing for the complete mineralization of the organic pollutant [
10,
11]. For this reason, bioremediation has been widely studied and is an environmentally friendly technology used for the removal of hydrocarbons in both terrestrial and aquatic ecosystems [
12]. The bioremediation of contaminated sites by organic compounds is based on the stimulation of the catabolic activity of microorganisms capable of using polluting organic contaminants as a source of carbon and energy. So far, bioremediation is performed through different practices such as biostimulation (the addition of macro- and/or micronutrients to enhance indigenous biomass growth and pollutant degradation), bioaugmentation (inoculation with pollutant-degrading microorganisms) or combined biostimulation and bioaugmentation [
13].
The microbial community in a given ecosystem is crucial for the biodegradation of pollutants to occur [
14]. In fact, bacterial adaptation, defined as an evolutionary process in which shifts in the microbial community composition or abundances take place in response to changes in environmental conditions and contaminant content, can improve the biodegradation rate of a chemical.
On the other hand, the existence of a microbial potential does not always lead to in situ biodegradation since many limitations, such as insufficient biomass, utilization of a wide range of substrates, competitive inhibition, or catabolite repression, can all inhibit the process [
14].
From this perspective, the analysis and characterization of microorganisms involved in biodegradation processes are of the utmost importance for the ultimate success of bioremediation. Furthermore, it is essential not only to dwell upon microbial communities but also have knowledge on the geological, hydrogeological and geochemical characteristics of the environment in which they live because several other factors may influence their activities [
5,
15].
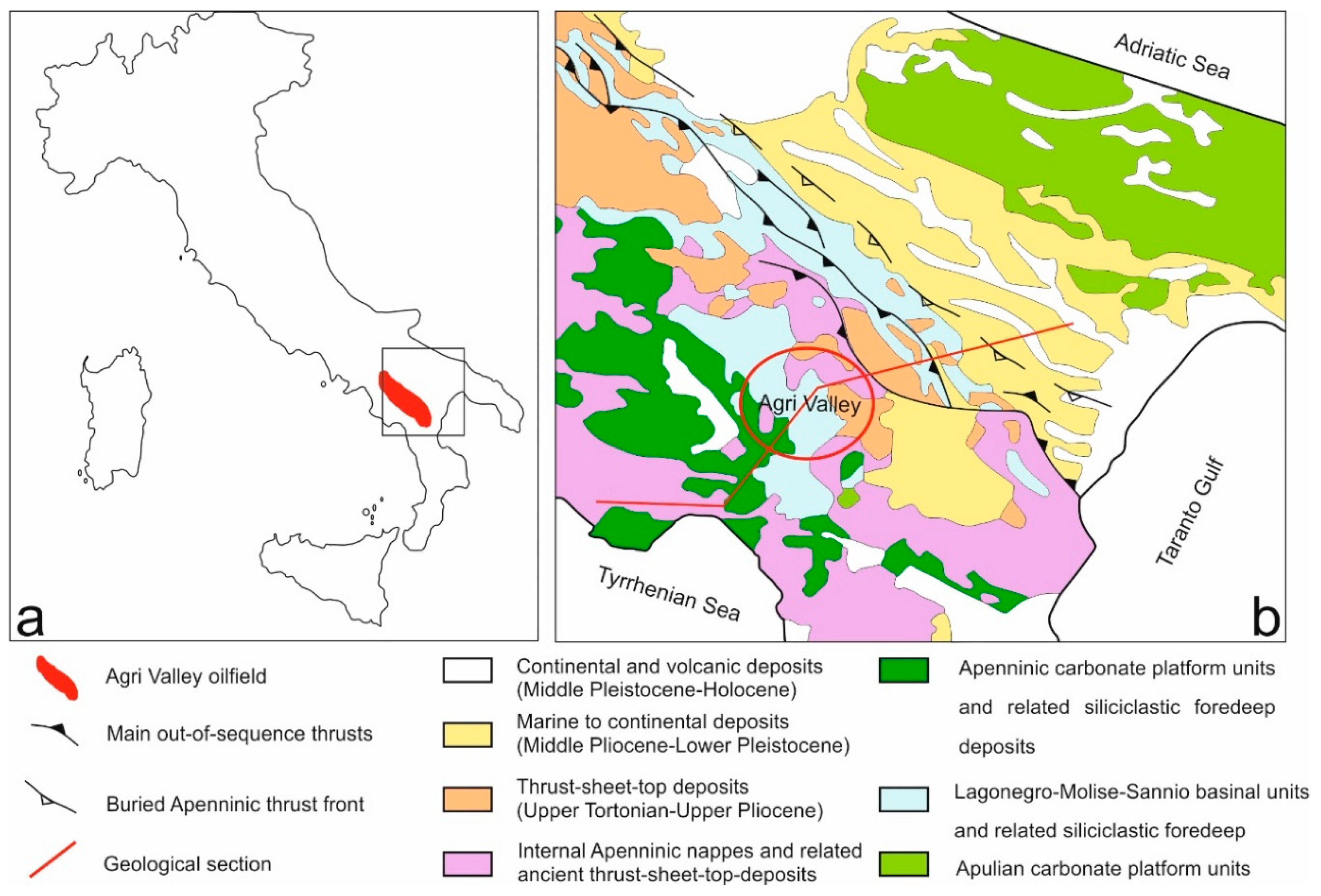
The main aim of the present work was to analyze microbial communities in soil, spring and groundwaters from a study site in southern Italy characterized by the presence of hydrocarbons of natural origin, through a combination of culture-dependent and molecular methods. In addition, the comprehension of the geological and hydrogeological features of the aquifer system was fundamental to identify the active circuits that continuously feed the studied soil, springs and groundwaters, all year round, and could influence the composition of the bacterial communities. Some of the bacterial isolates were identified and screened for their ability to grow in the presence of different pollutants and emulsifying capacity, to assess their potential as candidates in biotechnological applications to treat and recover the polluted environmental matrices.
4. Discussion and Conclusions
Petroleum hydrocarbon contamination is an issue of major concern worldwide. These compounds represent the most common environmental pollutants and their presence is destructive to the ecosystem and economic and human health [
44]. The clean-up of hydrocarbon-contaminated sites is expensive and time consuming; however, bioremediation is a cost-effective and environmentally safe approach for petroleum hydrocarbon contamination (PHC) removal [
44]. This technology involves the use of living organisms such as microbes and/or plants to reduce/degrade, eliminate and transform contaminants present in soils, sediments and water and has gained wider acceptance in recent years for all its potential [
45]. In fact, although oil pollution is difficult to treat, indigenous bacteria can ultimately degrade or metabolize most petroleum hydrocarbons encountered in the environment because of their energetic and carbon needs for growth and reproduction, as well as the requirement to relieve physiological stress caused by their presence [
46,
47,
48]. Accordingly, petroleum hydrocarbon-degrading bacteria, which have evolved as a result of existing in close proximity to naturally occurring petroleum hydrocarbons in the environment, represent suitable candidates for the treatment of oil polluted sites and to achieve the best purification effect [
49,
50,
51].
An understanding of the temporal and spatial structures, functions, interactions, and population dynamics of microbial communities is critical for biotechnological development, environmental protection, and human health [
52] so much that several methods have been employed to reveal microbial community composition, function and responses to environmental changes, in various environments and different contexts [
53,
54,
55,
56,
57,
58,
59,
60]. The main goal of this research was to analyze microbial communities in a site in southern Italy characterized by the presence of hydrocarbons of natural origin, by using a multidisciplinary approach based on microbiological, geological and hydrogeological investigations.
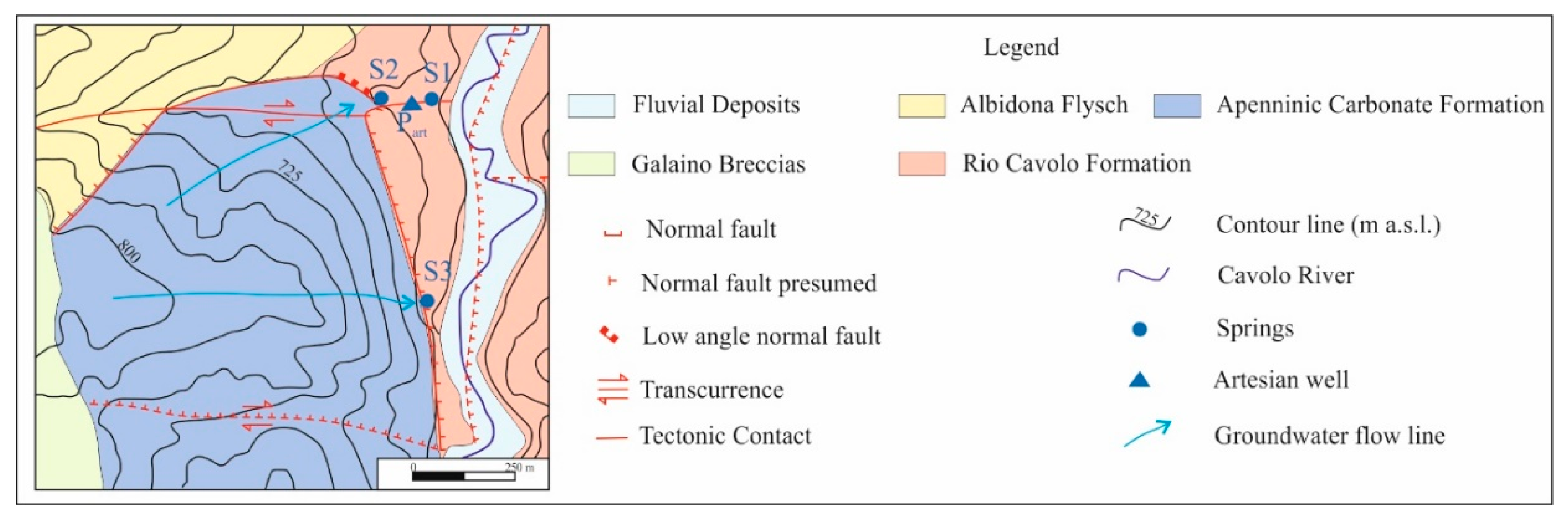
In detail, bacterial communities of two springs (S1 and S2), the surrounding soils, and groundwater (Part well) were studied through a combination of molecular and culture-dependent methodologies to explore the biodiversity at the study site, to isolate microorganisms with degradative abilities and to assess their potential to develop effective strategies to restore the environmental quality.
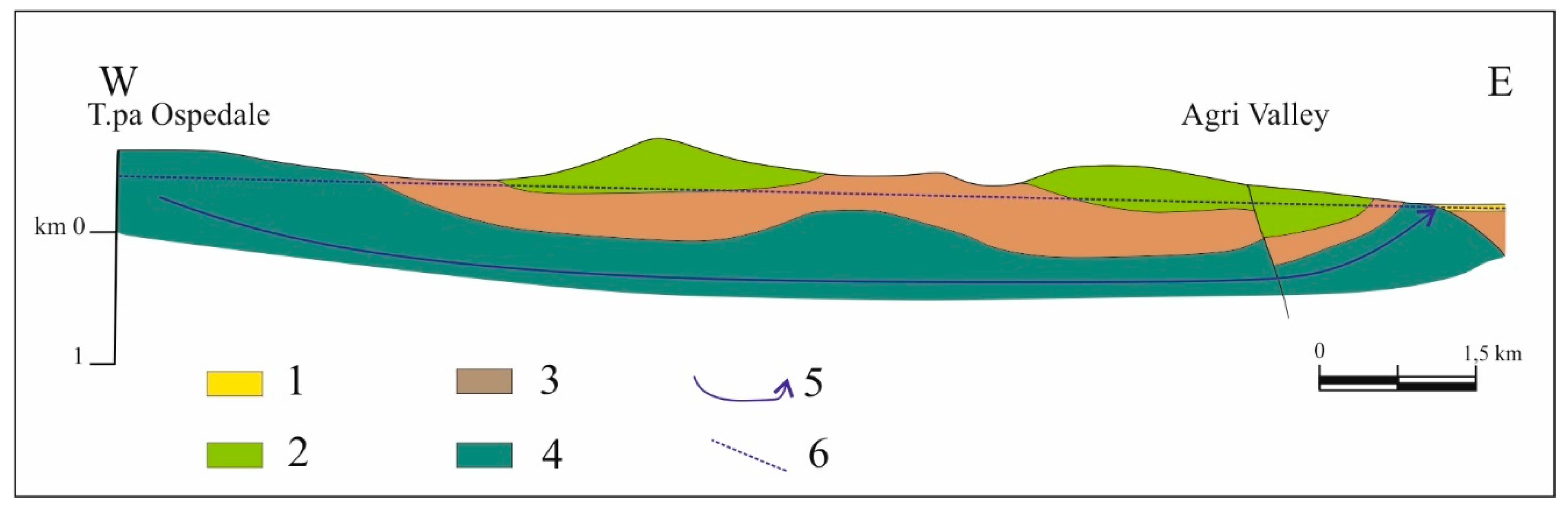
The two hydrocarbon springs S1 and S2 are linked to the recharge in a local aquifer system and distributed along a fault zone that favors the rise of fluids and hydrocarbons within the shallow low-permeability media. This hydrogeological behavior of the fault zone is not surprising when analyzing the possible roles (conduit, barrier, combined conduit-barrier systems) that these zones can play from the hydraulic point of view, depending on the relative percentage of fault core and damage zone [
61,
62,
63,
64,
65,
66,
67]. In many aquifer systems worldwide, comparable with the studied one, both structural and hydrogeological approaches demonstrated the possible migration of fluids within fault zones acting as high- or low-flow discontinuities [
68,
69,
70,
71,
72,
73,
74]. This evidence was further confirmed by the presence of H
2S at spring S1.
Chemical analyses of hydrocarbons highlighted the existence of several and different PAHs, such as Naphthalene, Benzo(b)fluoranthene, Benzo(k)fluoranthene, Benzo(g,h,i)perylene, Indene(1,2,3-c,d)pyrene, Chrysene, Anthracene, Phenanthrene, Fluoranthene, Benzo(j)fluoranthene, Dibenzo(a,h)anthracene, Pyrene and Dibenzo(a,l)pyrene in spring waters and soils. Generally, polycyclic aromatic hydrocarbons are chemicals with various structures and varied toxicity, stable, persistent in the environment, and resistant to degradation [
75,
76]. Chemically, they are comprised of two or more benzene rings bonded in linear, cluster, or angular arrangements [
75]. The increase in the number of fused rings leads to higher hydrophobicity and recalcitrance to microbial degradation [
77]. The continuous interaction with hydrocarbons could have influenced the bacterial communities of the two springs and the soils, naturally selecting some strains capable to biodegrade these pollutants.
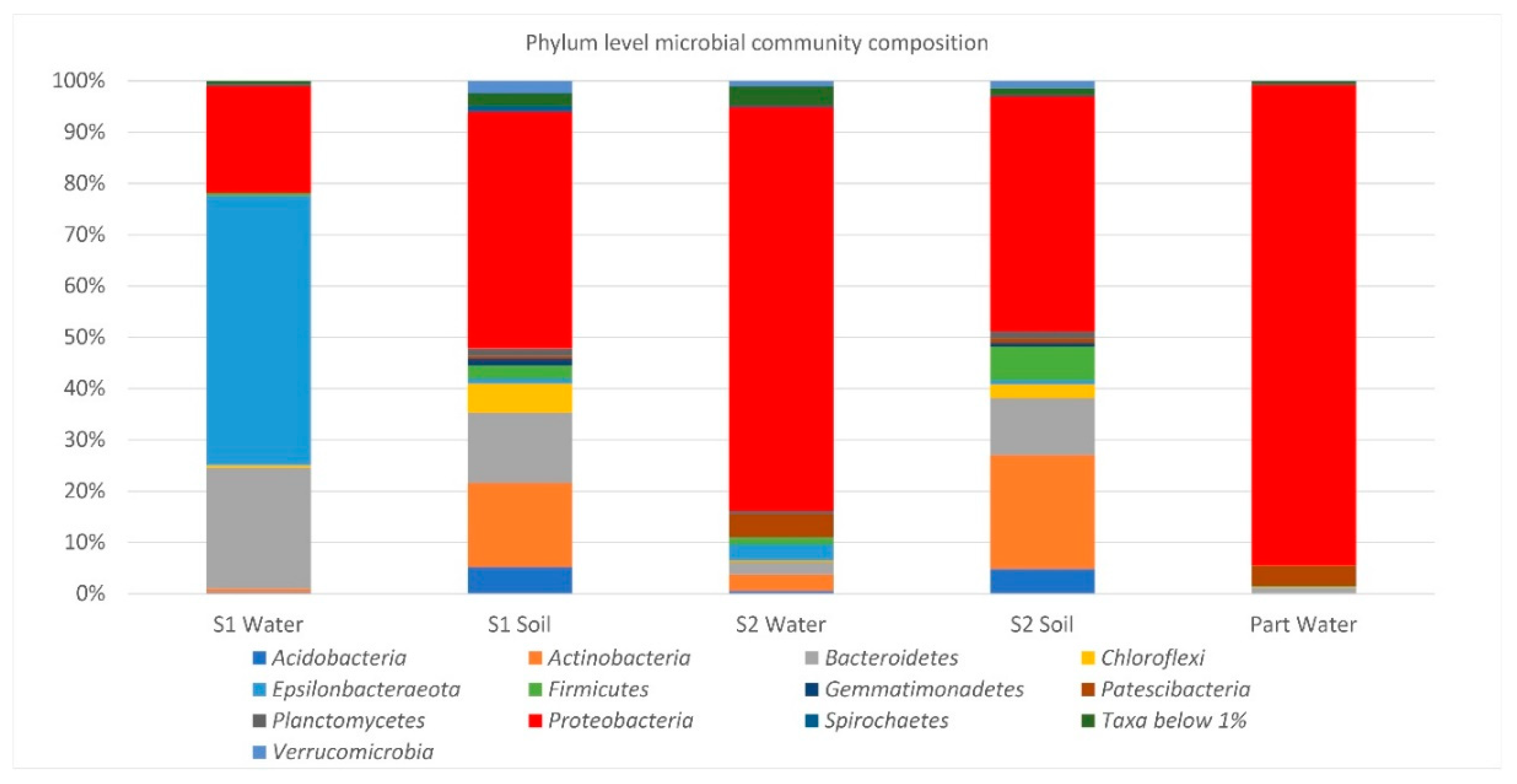
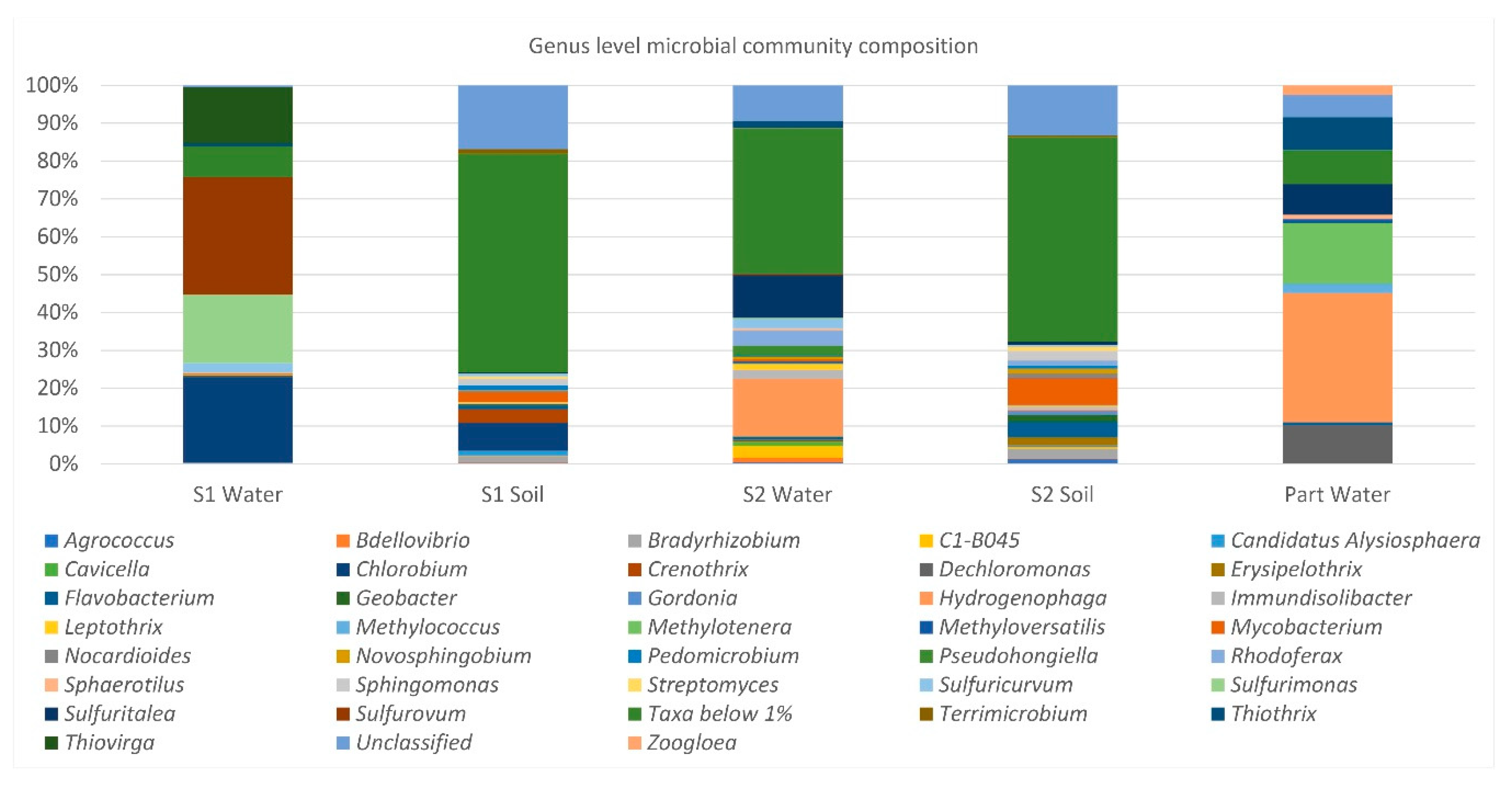
The NGS results allowed us to obtain detailed information about the composition of microbial communities and revealed the dominance of species belonging to the phylum
Proteobacteria. In this very large phylum of Gram-negative organisms, the majority of the formally described genera of hydrocarbon-degrading bacteria is included [
78]; thus, it is likely that the continuous outflow of oil over time led to a stable and highly specialized microbiota, particularly adapted to grow and thrive in those environmental conditions. However, in addition to
Proteobacteria, it was possible to also detect other autochthonous hydrocarbon-oxidizing bacteria affiliated to other phyla (e.g., species of the genera
Flavobacterium and
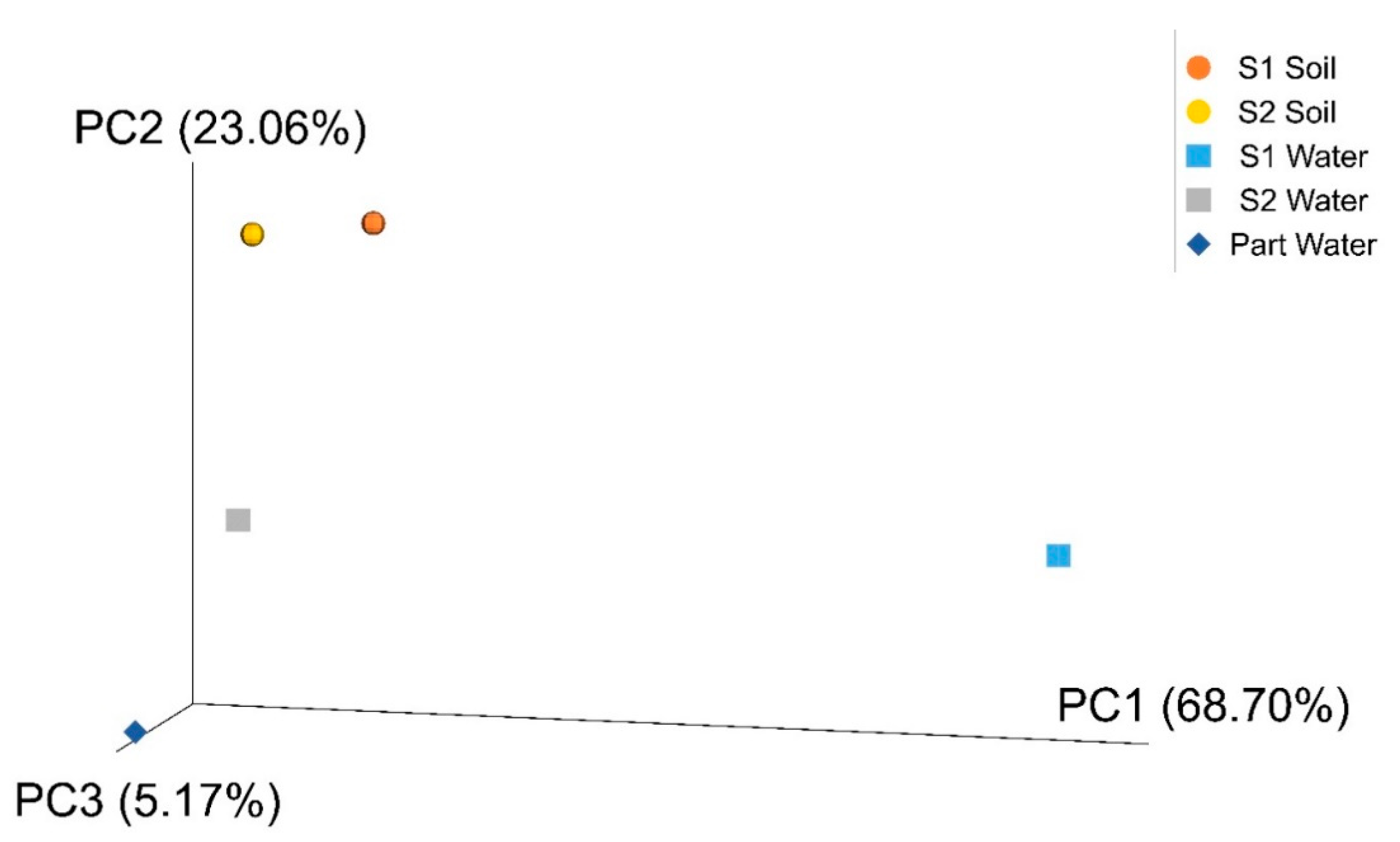
Gordonia), strengthening the assumption that natural exposure to pollutants can have impacted the structure and function of microbial populations. As expected, a higher biodiversity was found in soils compared to the aquatic ecosystems, generally lacking inputs of fresh, easily available organic carbon. In addition, beta-diversity analysis revealed a clear separation of soil, spring water, and groundwater samples and highlighted marked differences between S1 spring water microbial community and all the others. Although it is not surprising to find diverse microbial communities in different habitats due to their peculiar physico-chemical properties, it is likely that the observed results for the two springs reflect the influence of specific hydrogeological and other environmental factors that, on a local scale, shape bacterial community composition. As a matter of fact, P
art-groundwater is fed by a deep confined aquifer. Differently, the springs S1 and S2 are fed by a common groundwater coming from the nearby carbonate aquifer, mixed with different fluids (rising along a fault zone) in different proportions. This is clearly demonstrated by differences in some physico-chemical features (
Table 1) and hydrocarbon concentrations (
Table 2), and are further supported when taking into consideration the different mean residence time of spring waters, determined through tritium analyses (S1 = 5.6 to 6.9 tritium unit (TU), S2 = 6.8 to 7.5 TU; the spring fed by the only carbonate aquifer = 8.4 to 9.1 TU in Rizzo et al. [
19]). These results are also in agreement with findings in other hydrogeological settings, where differences in microbial communities were clearly explained through the differences in (i) the aquifer type, (ii) the mixing between shallow groundwater and ascending fluids, (iii) the mixing between waters characterized by different salinity [
53].
The traditional cultivation-based approach, performed by applying a selective pressure with diesel fuel supplementations, led to the isolation and identification of 11 aerobic hydrocarbon-oxidizing proteobacteria. Most of the bacteria isolated and identified belong, or are related, to genera or species known for their degradation abilities of polycyclic aromatic hydrocarbons [
79,
80,
81,
82,
83,
84,
85]. The results are in line with those obtained by chemical analyses, which showed higher concentrations of these compounds at the investigated site. It is interesting to note that microorganisms of the genera
Hydrogenophaga,
Sulfuritalea and
Sulfurovum, whose abundant presence was revealed by DNA-based analyses, have not been isolated after the enrichment procedures. This “paradox” can be explained by the theory of the “Great Plate Count Anomaly” [
86], which refers to the observation that only a small fraction (0.01–1%) of the microorganisms present in the environment can be cultivated in the laboratory. Despite this, the cultivation procedures led to the isolation of bacterial strains showing low values of 16S rRNA gene sequence identity with known species, contributing to get insights into the hidden microbial diversity of the analyzed ecosystem.
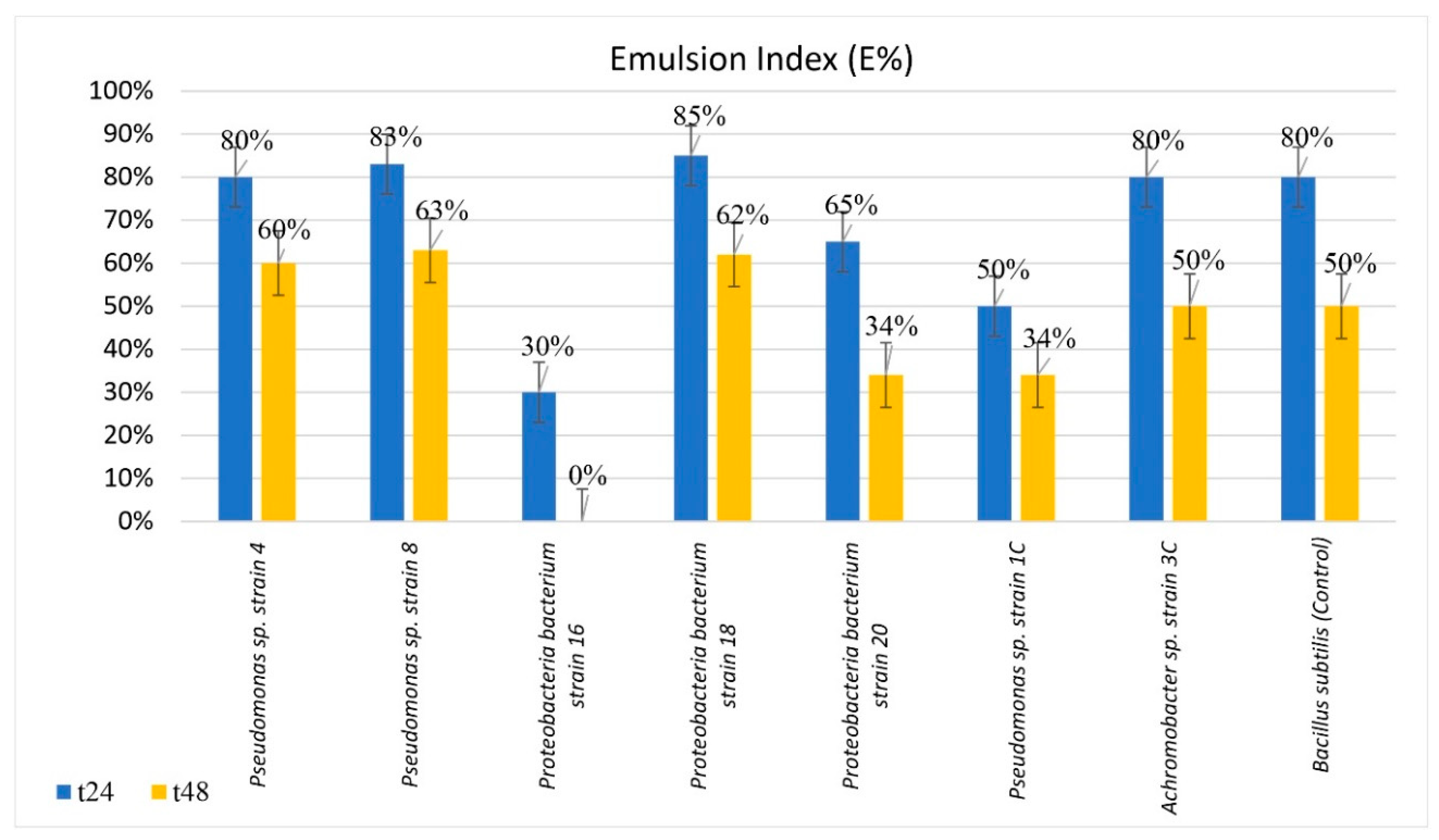
Some of the isolates were found to be able to grow better when phenanthrene was supplied as the sole carbon source compared to other simpler aliphatic and aromatic hydrocarbons. Subsequently, their emulsifying capacity was assessed and the emulsion index was calculated. Seven out of the 11 isolated bacterial strains produced emulsion with diesel fuel (most of them showing EC values greater than 50%) with high stability after 24 h and, in some cases, after 48 h. The tests have clearly demonstrated the capability of some strains to increase hydrocarbon bioavailability, most likely through the production of biosurfactants. These compounds are naturally derived surfactants produced from biological entities (especially microorganisms), which can be utilized as a cost-effective and eco-friendly mean to enhance bioremediation of oil components, including PAHs, in the natural environment. Fungi, bacteria, and yeasts belonging to different species and strains are known for producing biosurfactants of a diverse variety of molecular structures. Amongst the bacteria domain, the genera of
Pseudomonas,
Bacillus, and
Acinetobacter dominate the literature space as excellent producers of biosurfactants [
87]. For example,
Bacillus subtilis is known for its ability to enhance diesel solubility [
88] and to improve its degradation by producing surfactin, a lipoprotein-type biosurfactant [
89].
In the late 1960s, biosurfactants attracted attention as hydrocarbon-dissolving agents as potential replacements for synthetic surfactants (usually toxic, hardly degraded by microorganism, and causing damage to the environment) especially in the food, pharmaceutical, and oil industries [
90]. Bioemulsifiers get accumulated at the interphase between the two immiscible phases by which they can reduce the surface tension, thereby increasing the solubility and emulsification of the immiscible phases. Accordingly, these compounds may convert insoluble substrate into soluble substrates, increasing their bioavailability and making them usable by the microorganisms [
91].
In conclusion, the data collected in this study pave the way for further investigations finalized at exploiting both (i) the degradation ability of the bacterial isolates and/or microbial consortia to remediate hydrocarbon-contaminated sites through strategies like biostimulation or bioaugmentation, and (ii) the capability to produce molecules with a good promoting effect for the restoration of oil polluted matrices. In addition, the isolation of strains showing a 16S rRNA gene sequence identity <97% with those available in the genetic sequence database, constitutes a significant result and represents a further exciting challenge to fully unravel the existing biodiversity at the study site and expand the current knowledge on biodegradation processes, also through the description and characterization of novel bacterial species.

 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}